LET99에 의한 힘 발생기 공간 제한이 방추 위치를 견고하게 만든다
초록
이 연구는 C. elegans 초기 배아에서 방추의 비대칭 이동을 조절하는 두 가지 메커니즘을 규명한다. 하나는 세포 주기 진행에 따라 변화하는 힘 발생기의 결합·해리 역학(시간적 제어)이고, 다른 하나는 방추가 위치함에 따라 중심체와 세포 피질 사이의 미세소관 접촉 밀도가 변해 힘 발생기의 활성화를 조절하는 공간적 제어이다. LET‑99 단백질이 형성하는 후방 피질 밴드는 힘 발생기를 후방 초승달 영역에 제한함으로써 이 공간적 제어를 구현한다. 모델링과 실험을 결합한 결과, 이러한 이중 제어는 미세소관 수나 힘 발생기 수의 변동에 강인한 방추 위치를 보장한다는 것이 밝혀졌다.
상세 분석
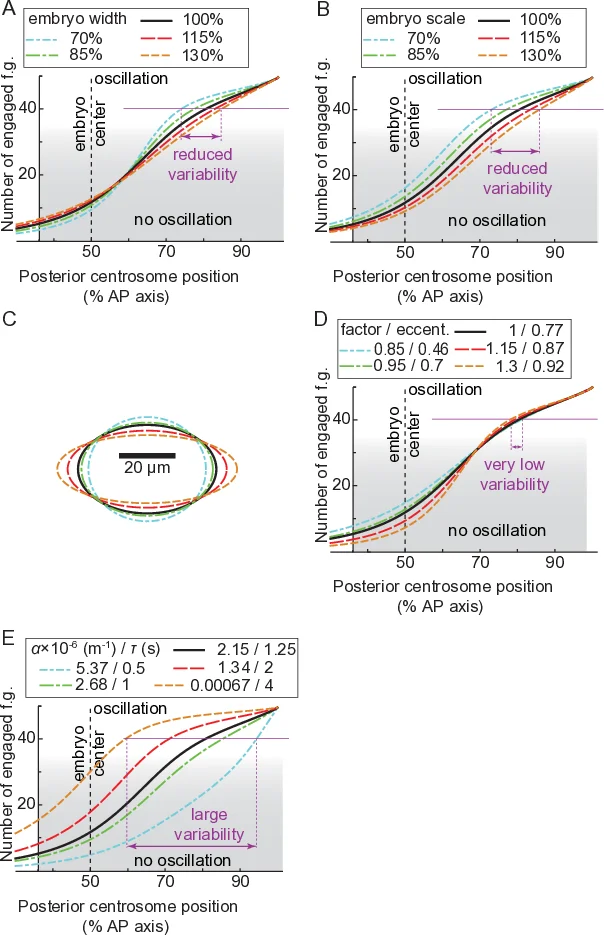
본 논문은 비대칭 세포 분열에서 방추 위치 결정 메커니즘을 정량적·정성적으로 해석한다. 기존 모델은 힘 발생기(FG)의 결합·해리 속도만을 조절 변수로 삼아, 전기적·화학적 신호가 미세소관 수에 비해 충분히 풍부하다고 가정하였다. 그러나 저자들은 미세소관이 피질에 닿는 확률이 중심체‑피질 거리(d)와 반비례한다는 실험적 근거를 제시한다. 이를 ‘landing assay’를 통해 10개의 등분된 AP 구간에서 접촉 밀도를 측정했으며, d가 짧아질수록 접촉 빈도가 급격히 증가함을 확인하였다. 이러한 거리 의존성은 미세소관 동역학(성장·소실 속도)과 피질에 존재하는 LET‑99 밴드가 결합한 FG의 공간적 분포가 결합될 때, 후방 초승달 영역에서만 유효한 FG가 활성화된다는 가설을 뒷받침한다.
시간적 제어는 APC/C 구성요소인 such‑1 돌연변이(ANAPC5)에서 관찰된 바와 같이, 무분열기 진행이 지연될 경우 방추 진동이 전형적인 무분열기 시점보다 앞서 발생한다는 사실로 입증된다. 이는 진동 시작이 순수히 세포 주기 타이밍에 의존하지 않고, 후방 중심체가 특정 위치(≈70 % EL, embryo length)까지 이동했을 때 자동으로 촉발된다는 것을 의미한다. 또한, SPD‑2(RNAi)로 미세소관 핵생성을 약화시키면 진동 시작 위치가 더 후방(≈74 % EL)으로 이동함으로써 미세소관 수 자체가 위치 감지에 기여함을 보여준다.
수학적 모델링은 두 가지 핵심 요소를 통합한다. 첫째, FG의 결합 확률은 미세소관 접촉 밀도 ρ(d)와 비례하고, ρ(d)는 실험적으로 추정된 지수 감소 함수이다. 둘째, LET‑99가 형성하는 피질 밴드는 FG의 활성화 영역을 45–70 % EL 구간에서 후방 70–100 % EL 구간으로 제한한다. 모델은 전후방 FG 수를 각각 Nₚ와 Nₐ로 두고, 결합·해리 속도 k_on, k_off을 시간에 따라 변화시키는 ‘프로세시비티’ 파라미터를 도입한다. 시뮬레이션 결과, 후방 FG가 제한된 영역에 집중될 경우, 미세소관 수가 20 % 감소하거나 FG 결합 속도가 30 % 변동해도 최종 방추 위치는 2–3 % EL 이내로 유지된다. 이는 ‘강인성(robustness)’이 미세소관·FG 수 자체보다는 공간적 제한 메커니즘에 의해 주도된다는 중요한 통찰을 제공한다.
또한, 저자들은 실험적 변이(embryo 길이 변화, 온도 변화)와 모델 예측을 비교함으로써, 후방 진동 시작 위치와 시점이 길이 정규화된 무차원 변수로서 거의 일정함을 확인하였다. 이는 세포 기계학적 피드백이 생화학적 네트워크보다 방추 위치를 안정화시키는 주된 원천임을 시사한다.
결론적으로, 이 연구는 (1) 미세소관-피질 거리 의존적 접촉 밀도, (2) LET‑99에 의한 힘 발생기 공간 제한, (3) APC/C‑의존적 프로세시비티 변화라는 세 축의 이중 제어가 비대칭 분열에서 방추 위치를 견고하게 만든다는 새로운 패러다임을 제시한다.
댓글 및 학술 토론
Loading comments...
의견 남기기