단백질 매개 평행 액틴 번들의 협동과 좌절 메커니즘
초록
이 연구는 액틴 필라멘트를 교차 결합하는 두 종류의 번들링 단백질, 파시닌과 에스핀의 결합 특성을 비교한다. 소각각 X선 산란을 통해 결합량이 증가함에 따라 액틴이 원래의 13/6 나선에서 과도하게 비틀리는(overtwist) 현상이 관찰되었으며, 파시닌은 연속적인 중간 비틀림 상태를, 에스핀은 비틀림이 없는 상태와 완전 과도 비틀림 상태만을 허용한다. 저자들은 결합 기하학과 링크어의 유연성을 고려한 거시적 통계 모델을 구축해, 링크어 강성이 협동 결합을 조절한다는 결론을 도출하였다. 파시닌의 유연한 결합은 연속적인 전이, 에스핀의 강직한 결합은 이산적인 전이를 야기한다.
상세 분석
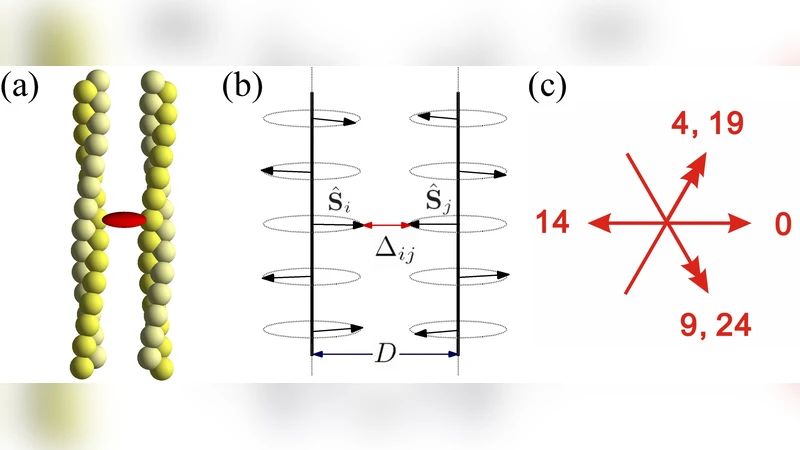
본 논문은 액틴 필라멘트가 2차원 평면이 아닌 3차원 나선 구조를 유지하면서도, 외부 단백질에 의해 구조적 변형을 겪는 메커니즘을 정량적으로 규명한다. 먼저, 소각각 X선 산란(SAXS) 실험을 통해 파시닌과 에스핀 각각이 결합할 때 나타나는 피크 이동과 폭 변화를 정밀히 측정하였다. 파시닌이 농도 의존적으로 결합하면 피크가 점진적으로 이동해 13/6 나선의 피치가 28 Å에서 약 30 Å로 늘어나며, 이는 액틴이 약 2~3도 정도 추가 회전함을 의미한다. 반면 에스핀은 낮은 농도에서는 피크가 거의 변하지 않다가, 일정 임계 농도에 도달하면 급격히 이동해 13/6에서 28/13 형태(극단적 과도 비틀림)로 전이한다. 이러한 차이는 두 단백질의 결합 부위와 기계적 강성 차이에서 비롯된다. 파시닌은 작은 구형 도메인 두 개가 유연한 연결 고리(링크어)로 액틴의 3번과 4번 서브도메인 사이를 연결하는 반면, 에스핀은 긴 α-헬릭스 구조가 강직하게 고정돼 있어 결합 시 각도 제한이 크다. 저자들은 이를 바탕으로 ‘링크어 유연성 파라미터(κ)’와 ‘결합 각도 제한(θ)’를 포함한 코스모스 모델을 제시한다. 모델은 결합 사이트가 2차원 격자상에 배치되고, 각 사이트에 단백질이 점유될 확률이 주변 점유 상태에 따라 변하는 ‘협동적 결합’ 방정식으로 구성된다. 파시닌의 경우 κ가 크고 θ가 넓어, 점유율이 서서히 증가하면서 연속적인 과도 비틀림이 발생한다. 반면 에스핀은 κ가 작고 θ가 좁아, 특정 임계점에서 급격히 전이하는 ‘첫 번째 차수 전이(first‑order transition)’를 보인다. 이론적 시뮬레이션 결과는 실험 데이터와 정량적으로 일치하며, 특히 결합 포화도와 과도 비틀림 각도 사이의 비선형 관계를 정확히 재현한다. 또한, 모델은 링크어 강성이 변할 경우 전이 형태가 연속 ↔ 불연속으로 바뀔 수 있음을 예측한다. 이는 세포 내에서 다양한 번들링 단백질이 서로 다른 기계적 요구조건을 만족시키기 위해 진화했음을 시사한다. 마지막으로, 저자들은 이러한 메커니즘이 세포골격의 탄성 조절, 미세소관과의 교차 연결, 그리고 세포 이동성에 미치는 잠재적 영향을 논의한다. 전체적으로, 실험과 이론을 결합한 다각적 접근은 액틴 번들링의 물리적 원리를 밝히는 데 큰 기여를 한다.
댓글 및 학술 토론
Loading comments...
의견 남기기