무한주기 분기 현상이 세분화 시계 정지 메커니즘을 밝히다
초록
본 논문은 척추동물의 체절 형성 과정에서 중요한 역할을 하는 세분화 시계(분절 시계)의 진동이 어떻게 멈추는지를 최소 모델링을 통해 규명한다. Wnt, Notch, FGF 세 경로와 RA‑Fgf8 상호작용을 포함한 네트워크 구조를 단순화하고, Wnt3a 구배를 위치 신호(k)로 도입해 분석하였다. 결과는 k가 특정 임계값 이하로 감소할 때 무한주기(SNIC) 분기가 발생해 진동이 서서히 주기가 길어지며 진폭이 커진 뒤 고정된 높은 활성 상태로 전이된다는 것을 보여준다. 전이 과정은 히스테리시스를 동반하며, 모델은 실험적으로 관찰된 앞쪽 PSM에서의 진동 감속·증폭·정지 현상을 재현한다.
상세 분석
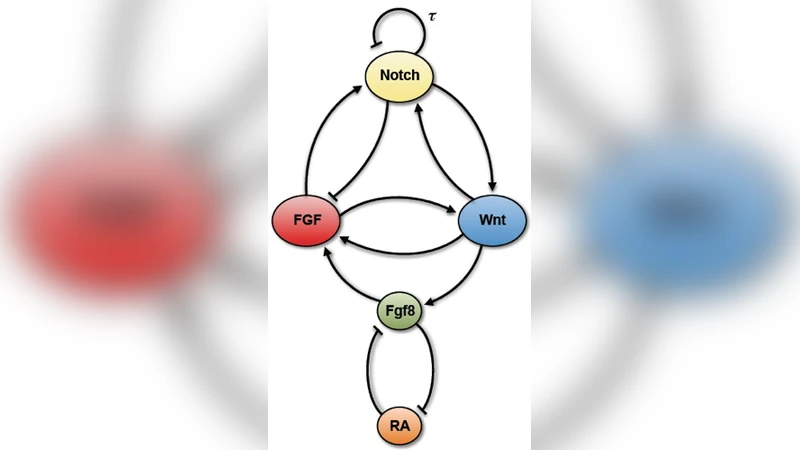
이 연구는 복잡한 amniote(포유류·조류) 체절 형성 네트워크를 “구조‑우선” 접근법으로 단순화한다. 핵심은 세 가지 신호 경로(Wnt, Notch, FGF)를 각각 하나의 변수(w, n, f)로 요약하고, RA와 Fgf8을 x, y로 표현한 5차 지연 미분 방정식이다. Notch 경로는 지연된 음성 피드백(τ≈45 min)으로 자체 진동을 생성하고, Wnt‑Notch 사이의 양성 상호작용은 이 진동에 대한 bistable 스위치를 제공한다. 또한 Wnt‑FGF 양성 연결과 Notch‑FGF 음성 연결을 포함해 전체 네트워크가 상호 얽히게 된다. 파라미터 β와 K는 Hill 함수 형태(계수 4)로 비선형성을 부여했으며, 특히 β_wn, β_nw를 크게 설정해 Wnt‑Notch 교차 조절을 지배적인 양성 회로로 만든 점이 핵심이다.
위치 정보는 Wnt3a 구배를 k(1→0)로 매핑함으로써 두 가지 효과를 동시에 구현한다. 첫째, k가 감소하면 Notch 수용체 NICD의 분해율 γ_n이 증가해 Notch 진동이 약화된다. 둘째, Wnt‑FGF 양성 연결 β_xw가 감소해 Fgf8 생산이 억제되고, 결과적으로 RA와 Fgf8 사이의 상호 억제 회로가 전방에서 RA가 높고 후방에서 Fgf8이 높은 반대 구배를 형성한다.
bifurcation 분석 결과, k≈0.44에서 SNIC(무한주기) 분기가 발생한다. 이 분기 전에는 안정적인 한계 주기가 존재하지만, k가 감소함에 따라 주기가 점점 길어지고 진폭이 커진다(‘slow‑down’ 현상). 임계값 이하에서는 진동이 사라지고 시스템은 높은 Wnt·Notch 활성과 중간 수준의 FGF 활성으로 고정된다. 흥미롭게도, k를 다시 증가시켜도 진동이 재개되는 임계값은 k≈0.76으로, 큰 차이의 히스테리시스 구역이 형성된다. 이는 실험적으로 보고된 ‘irreversible’ 체절 전환과 일치한다.
모델은 또한 Fgf8/RA 구배가 자연스럽게 반대 방향으로 형성되는 메커니즘을 제공한다. Wnt3a 감소가 직접 Fgf8 전사를 억제하고, 동시에 RA는 Fgf8을 억제하는 이중 부정 회로를 통해 앞쪽 PSM에서 RA가 축적되고, 뒤쪽에서는 Fgf8이 우세하게 된다. 이러한 결과는 기존의 Fgf8‑RA bistable 도메인 가설을 수학적으로 뒷받침한다.
전반적으로, 이 논문은 “진동을 일으키는 Notch 지연‑음성 피드백 + Wnt‑Notch 양성 교차 회로”라는 최소 구조가 amniote 체절 시계의 핵심 동역학을 설명한다는 강력한 증거를 제시한다. 모델은 복잡한 분자 세부사항을 배제하고도 실험적 관찰(진동 감속, 진폭 증가, 히스테리시스, 구배 형성)을 정량적으로 재현한다는 점에서, 구조적 접근법의 가치를 잘 보여준다.
댓글 및 학술 토론
Loading comments...
의견 남기기