클러스터 기반 집합·분해 모델로 구현한 세포 내 신호 농도 구배의 강인성
초록
본 논문은 베커‑도링(Bekker‑Döring) 방정식을 활용해 단일 분자 클러스터가 형성·분해되는 과정을 모델링하고, 이러한 클러스터 매개 메커니즘이 세포 내 신호 농도 구배를 시간·공간적으로 실현 가능한 범위 내에서 강인하게 유지할 수 있음을 보인다. 특히 클러스터가 신호 분자의 해리 속도를 크게 증가시키지 않을 때, 그리고 삽입·집합·분해·확산 파라미터가 실험값과 일치할 때, 모델은 fission yeast의 pom1p 구배와 정량적으로 일치한다.
상세 분석
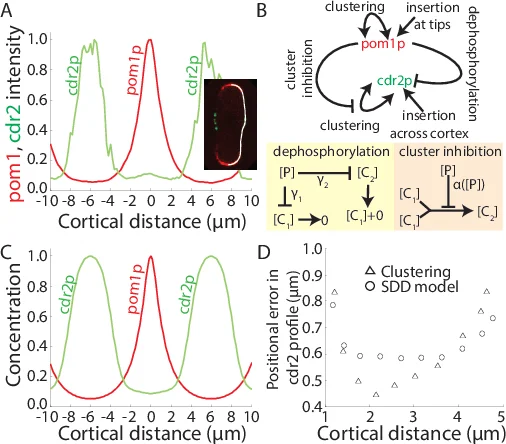
본 연구는 세포 내 신호 분자(예: pom1p)의 클러스터링 현상을 물리‑수학적으로 정량화하기 위해 베커‑도링(Bekker‑Döring) 형태의 집합·분해 방정식을 채택하였다. 핵심 가정은 (1) 클러스터는 단일 모노머가 하나씩 흡수·방출되는 방식으로 성장·소멸한다는 점, (2) 클러스터 자체가 막에서 동시에 탈착되는 현상은 관찰되지 않아 무시한다는 점, (3) 삽입은 오직 모노머 형태로만 일어난다. 이러한 가정은 pom1p 실험 영상에서 관찰된 “클러스터가 독립적으로 성장·소멸한다”는 현상을 충실히 반영한다.
수식(1‑3)은 각각 모노머와 s‑크기 클러스터의 농도 변화를 기술한다. 여기서 확산 계수 D_s는 D_1·s^(-r_D) (r_D≈1) 로 스케일링하고, 해리율 μ_s는 μ_1·s^(-r_μ) 로 정의한다. r_μ=0이면 클러스터가 신호 분자의 안정성을 변화시키지 않으며, r_μ=1이면 클러스터 내부에서 분자가 더 오래 머무른다(안정화). 저자는 r_μ=0인 경우에만 구배가 강인함을 보이며, 이는 클러스터가 효과적인 확산을 조절하면서도 전체 신호 수명을 크게 늘리지 않기 때문이다.
파라미터 차원 축소를 통해 7개의 무차원 변수(κ, β̃ 등)만이 동역학을 지배함을 증명한다. κ=J·α_1/(√(D_1)·μ_1^3) 은 삽입·집합·확산·해리의 상대 비율을 나타내며, β̃=β_1/μ_1 은 클러스터 내 분해 속도를 조절한다. 실험값을 대입하면 κ≈425, β̃≈20 정도가 얻어지며, 이는 클러스터가 충분히 형성되면서도 과도히 큰 클러스터가 발생하지 않는 ‘중간’ 영역에 해당한다.
수치 해석(pde45, bvp4c) 결과, 총 농도 N_T(x)=∑_s s·n_s(x)는 소스 근처에서 급격히 감소하고, 멀리 갈수록 평탄해진다. 클러스터 크기별 농도 분포는 큰 클러스터가 소스에 국한되고, 작은 클러스터가 멀리 퍼지는 형태를 보인다. 효과적 확산 D_eff(x)=∑_s s·n_s·D_s / N_T는 위치에 따라 증가하는데, 이는 클러스터가 고농도 영역에서 확산을 억제하고 저농도 영역에서 확산을 촉진함을 의미한다. 따라서 클러스터링은 ‘확산 조절 메커니즘’으로 작동한다.
강인성 분석에서는 (i) 삽입율 J 변동에 대한 위치 오차 δx, (ii) 내재적(포아송) 잡음에 대한 δx를 평가한다. 기존의 선형 확산·분해(SDD) 모델과 비선형 분해(NLD) 모델을 비교했을 때, 클러스터링 모델은 J 변동에 대해 NLD와 동등하거나 더 작은 δx를 보이며, 내재적 잡음에 대해서는 CDD(농도‑의존 확산) 모델보다 약간 떨어지지만 여전히 실험적 허용 범위 내에 있다. 특히 r_μ=0인 경우에만 이러한 강인성이 유지되며, r_μ=1이면 클러스터가 신호를 과도히 안정화시켜 구배가 급격히 변하고 민감도가 증가한다.
마지막으로 pom1p 구배에 모델을 적용했을 때, 실험에서 보고된 클러스터 크기 분포와 농도 프로파일이 정량적으로 일치한다. 이는 모델이 실제 세포 내 신호 전달 메커니즘을 포착하고 있음을 강력히 시사한다. 전체적으로 본 연구는 클러스터링이 단순한 ‘신호 증폭’이 아니라, 공간적 확산 조절과 시간적 안정성을 동시에 제공하는 복합 메커니즘임을 증명한다.
댓글 및 학술 토론
Loading comments...
의견 남기기