리듬을 만드는 신경망과 k‑코어 구조
초록
본 논문은 호흡 리듬을 담당하는 전뇌교(pre‑Bötzinger) 복합체를 최소 모델링하여, 동일한 흥분성 뉴런이 무작위로 연결된 경우 네트워크 전체의 발화 주기가 어떻게 emergent pacemaker와 k‑코어 클러스터에 의해 조절되는지를 분석한다. 평균장 이론과 달리 연결 구조가 발화 시작과 종료 시점에 결정적인 역할을 함을 보여준다.
상세 분석
이 연구는 호흡 조절에 핵심적인 전뇌교(pre‑Bötzinger complex, preBötC)의 동적 메커니즘을 이해하기 위해, 동일한 흥분성 뉴런들로 이루어진 무작위 연결 네트워크를 수학적으로 구현하였다. 기존의 평균장(mean‑field) 접근법은 모든 뉴런이 동일한 평균 입력을 받는다고 가정해, 네트워크 전체가 동기화된 발화와 소멸을 일괄적으로 설명한다. 그러나 실제 생물학적 네트워크는 연결도가 이질적이며, 특정 뉴런이 다른 뉴런보다 더 많은 시냅스 입력을 받는다. 저자들은 이러한 이질성을 반영하기 위해, 각 뉴런의 입출력 관계를 단순한 발화‑재활성화 모델로 설정하고, 연결 행렬을 에르되시‑레니(Erdős‑Renyi) 방식의 확률적 그래프 형태로 생성하였다.
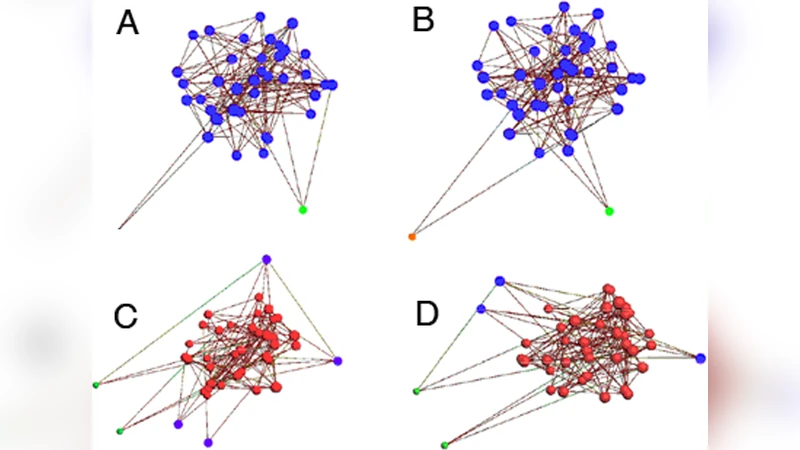
핵심 발견은 두 가지이다. 첫째, 네트워크 내에서 연결도가 상대적으로 높은 ‘핵심’ 뉴런들이 자연스럽게 emergent pacemaker 역할을 수행한다는 점이다. 이 뉴런들은 다른 뉴런들보다 빠르게 탈분극에 도달하고, 그 결과 전체 네트워크에 파동 형태의 발화를 전파한다. 이러한 pacemaker는 사전에 지정된 것이 아니라, 연결 구조에 의해 스스로 선택된다. 둘째, 발화가 지속되는 동안 뉴런들은 점진적으로 탈감작(desensitization)되어 활동 전압을 감소시키는데, 이 과정은 네트워크의 k‑core 구조와 밀접하게 연관된다. k‑core는 그래프에서 각 노드가 최소 k개의 연결을 유지하는 최대 서브그래프를 의미한다. 저자들은 발화 종료 시점에 네트워크가 특정 k‑core(예: k=3 또는 k=4) 이하로 붕괴되는 현상을 관찰했으며, 이는 충분히 높은 차수의 연결을 가진 클러스터가 사라질 때 전체 네트워크가 동기화된 발화를 멈추는 메커니즘으로 해석한다.
수치 시뮬레이션 결과는 평균장 모델이 예측하는 임계 연결도와 실제 무작위 네트워크에서 관찰되는 임계값 사이에 큰 차이가 있음을 보여준다. 특히, 연결도 p가 평균장 이론에서 제시하는 최소값보다 낮아도, 특정 고차수 노드가 존재하면 발화가 가능하고, 반대로 p가 충분히 높아도 k‑core가 충분히 강건하지 않으면 발화가 지속되지 않는다. 이는 네트워크 토폴로지가 동적 특성을 결정하는 주요 변수임을 강력히 시사한다.
이러한 결과는 신경생리학적 실험과도 일맥상통한다. 전뇌교 내에서 특정 ‘핵심’ 뉴런이 광유전학적 자극에 의해 발화 시작을 주도한다는 보고와, 네트워크 전체가 일정 시간 후 자동으로 억제되는 현상이 관찰된 바 있다. 따라서 이 논문의 모델은 실험적 관찰을 이론적으로 뒷받침하며, 향후 약물 표적이나 전기 자극을 통한 호흡 리듬 조절 전략을 설계하는 데 중요한 지침을 제공한다.
요약하면, 무작위 연결된 동일 흥분성 뉴런 네트워크는 평균장 접근법으로는 설명할 수 없는 복합적인 발화 메커니즘을 보이며, 연결도 분포에 의해 emergent pacemaker와 k‑core 기반 탈감작이 동시에 작동한다. 이는 신경망 연구에서 토폴로지와 동역학의 상호작용을 고려해야 함을 강조한다.
댓글 및 학술 토론
Loading comments...
의견 남기기