단백질 2차 구조와 결합 결합 대칭 파괴
초록
본 연구는 단백질 골격의 Cα 원자 주변에 존재하는 sp³ 하이브리드 탄소의 정사면체 대칭이 실제 활성 단백질에서 구조에 따라 체계적으로 깨진다는 사실을 고해상도 PDB 데이터를 통해 입증한다. 특히 N‑Cα‑C 결합각(τ_NC)이 α‑헬릭스, β‑시트, 3/10‑헬릭스 등 2차 구조에 따라 변동함을 확인하고, 이를 설명하기 위해 이산 비선형 슈뢰딩거(DNLS) 방정식 기반의 새로운 에너지 함수와 솔리톤 해를 제시한다. 실험적 B‑factor 수준의 RMSD(0.39 Å) 정확도로 Cβ 위치까지 재현함으로써, 기존의 무대칭 가정이 과도하게 단순화된 것을 지적한다.
상세 분석
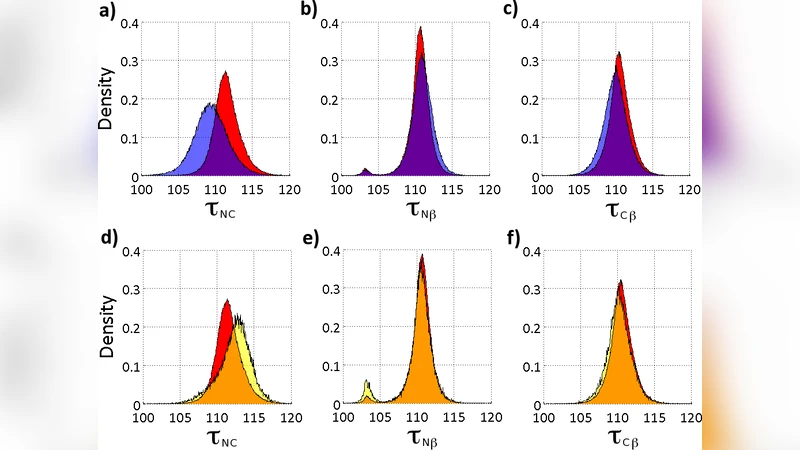
이 논문은 먼저 전통적인 단백질 모델링이 전제하는 “Cα 탄소는 완전한 sp³ 정사면체 구조를 유지한다”는 가정을 검증한다. 최신 2 Å 이하 해상도의 X‑ray 구조 데이터를 이용해 τ_NC, τ_Nβ, τ_Cβ 세 종류의 결합각을 각각 α‑헬릭스, β‑시트, 3/10‑헬릭스 구간에서 통계적으로 분석했으며, τ_NC만이 2차 구조에 따라 뚜렷한 변화를 보이는 것을 발견했다. 이는 N‑Cα 결합이 주변 전자 구름 및 수소 결합 네트워크에 의해 구조적으로 편향된다는 의미이며, 기존 힘장 모델이 가정하는 일정한 평형각(θ₀)이 실제 단백질에서는 부정확함을 시사한다.
다음으로 저자들은 프레넷 프레임을 이용해 백본을 기하학적으로 기술하고, 각 잔기의 탄소‑탄소 결합각 κ_i와 비틀림각 τ_i를 DNLS 방정식의 해석 변수로 전환한다. DNLS는 보존량이 무한히 존재하는 적분가능계이며, 여기서 제시된 에너지 함수(식 4)는 κ_i와 τ_i의 2차항, 그리고 τ_i와 κ_i² τ_i와 같은 저차 보존량을 포함한다. 특히 τ_i가 2차 이하로만 나타나므로, τ_i는 κ_i에 대한 방정식으로부터 명시적으로 제거 가능하고, 결과적으로 백본의 기하학은 κ_i 솔리톤 프로파일에 의해 완전히 결정된다.
그 후 저자들은 위의 이론을 실제 단백질인 HP35(1YRF) 에 적용하였다. 두 개의 솔리톤을 이용해 루프 구간을 모델링하고, 식 5·6을 통해 θ_i와 φ_i(즉, Cβ의 구면 좌표)를 κ_i에 의존하도록 계산하였다. 파라미터 최적화 후 얻어진 RMSD는 Cα와 Cβ 모두 0.38 Å 이하로, 실험 B‑factor 수준의 정확도를 달성했다. 이는 기존의 “정사면체 대칭 유지” 가정이 불필요함을 실증적으로 보여준다.
논문은 또한 대칭 파괴가 단순히 구조적 편향이 아니라, 단백질이 폴딩될 때 에너지 지형의 최소화 과정에서 자연스럽게 발생하는 현상임을 강조한다. DNLS 솔리톤이 제공하는 비선형, 비국소적 상호작용은 폴딩된 단백질의 전역적인 콜랩스 현상을 설명하며, 이는 기존의 조화적 근사(force field)와는 근본적으로 다른 메커니즘이다. 마지막으로 저자들은 초고해상도 X‑ray 실험과 양자화학 계산을 통해 τ_NC의 구조 의존성을 직접 검증할 필요성을 제시하고, 향후 약물 설계 및 단백질 설계에 이 새로운 에너지 모델이 활용될 가능성을 논의한다.
댓글 및 학술 토론
Loading comments...
의견 남기기