전하 코어 주변 바이러스 캡시드 조립 이론: 임계 전하밀도와 동역학적 특성
초록
이 논문은 전하를 띤 나노입자(폴리전해질로 기능화) 주변에 바람꽃 바이러스(BMV) 캡시드 단백질이 스스로 조립되는 과정을, 평형 및 동역학 모델을 통해 설명한다. 비선형 포아송‑볼츠만 방정식으로 폴리전해질 브러시와 반대 전하 표면 간 상호작용을 계산하고, 기능화된 전하밀도가 일정 임계값을 초과하면 캡시드가 효율적으로 입자를 감싸며, 조립 초기 단계에서 빛 산란 강도가 급격히 증가하고 빈 캡시드 조립에서 보이는 지연 단계가 사라진다는 예측을 제시한다. 실험 데이터와 전반적으로 일치하지만, 전하밀도 의존성이 과도하게 강하고, 로그 성장 단계가 완전히 재현되지 않아 메타안정적 무질서 상태의 존재를 시사한다.
상세 분석
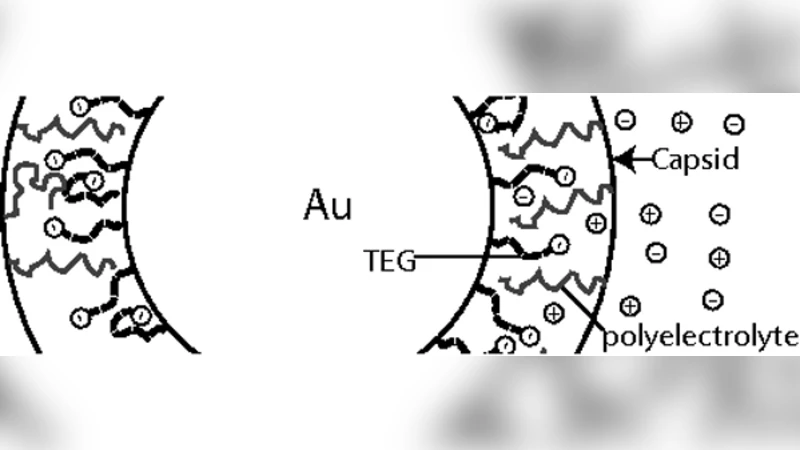
본 연구는 바이러스 캡시드 단백질이 전하를 띤 핵심(core) 주변에서 자가조립되는 현상을 정량적으로 이해하기 위해 두 가지 이론적 틀을 구축한다. 첫 번째는 열역학적 평형 모델로, 캡시드 단백질과 나노입자 표면 사이의 전기적 상호작용을 ‘폴리전해질 브러시‑반대 전하 표면’ 시스템으로 환원한다. 비선형 포아송‑볼츠만 방정식을 수치적으로 풀어 전위와 이온 분포를 얻고, 이를 통해 전하밀도(σ)와 입자 반경(R) 사이의 자유에너지 함수를 도출한다. 이 자유에너지 곡선은 σ가 특정 임계값 σ_c를 초과할 때, 캡시드가 입자를 포위하는 ‘핵심‑캡시드 복합체’가 전역 최소가 됨을 보여준다. 즉, 전하밀도가 충분히 높으면 전기적 인력이 캡시드-캡시드 간의 반발을 압도해 조립을 촉진한다는 것이다.
두 번째는 동역학 모델로, 캡시드 단백질이 자유 상태에서 핵심에 결합하고, 이후 서로 결합해 완전한 icosahedral 구조를 이루는 과정을 마스터 방정식 형태로 기술한다. 핵심‑단백질 결합 속도(k_on)와 해리 속도(k_off)는 전위에 의존하는 Boltzmann 인자 exp(−ΔG_elec/k_BT) 로 조정되며, 캡시드‑캡시드 결합은 전형적인 ‘핵심‑촉진’ nucleation‑growth 메커니즘을 따른다. 모델은 초기 단계에서 핵심에 단백질이 빠르게 흡착되면서 빛 산란 강도가 급격히 상승하고, 전통적인 빈 캡시드 조립에서 관찰되는 ‘lag phase’가 사라지는 현상을 재현한다. 이는 핵심이 ‘핵’ 역할을 수행해 핵생성 장벽을 실질적으로 없애기 때문이다.
하지만 실험과 비교했을 때 두 가지 중요한 차이가 드러난다. 첫째, 실험에서는 전하밀도 σ가 증가함에 따라 캡시드 포장 효율이 완만하게 상승하지만, 모델은 σ가 σ_c 근처에서 급격한 전이(첫 번째 오더 파라미터 급증)를 예측한다. 이는 모델이 전하 분포를 평균화하고, 브러시‑브러시 상호작용을 단순화함으로써 실제 폴리전해질 사슬의 유연성 및 공간적 이질성을 충분히 반영하지 못했기 때문일 가능성이 있다. 둘째, 실험에서 관찰된 로그 성장 단계(빛 산란이 시간에 따라 로그 형태로 증가)는 현재의 마스터 방정식이 포착하지 못한다. 저자들은 이를 ‘메타안정적 무질서 상태’가 존재해, 캡시드가 완전한 icosahedral 형태로 전환되기 전 일시적인 비정질 집합체를 형성한다는 가설을 제시한다. 이러한 상태는 에너지 장벽이 낮아 재배열이 느리게 진행되면서 로그 형태의 성장 곡선을 만든다.
결론적으로, 이 논문은 전하 코어 주변 캡시드 조립을 설명하는 첫 번째 정량적 프레임워크를 제공한다. 전하밀도 임계값, 초기 빛 산란 급증, lag phase 소멸 등 핵심적인 현상을 성공적으로 설명하지만, 전하밀도 의존성 과대평가와 로그 성장 미포착이라는 한계가 있다. 향후 모델에 폴리전해질 사슬의 구조적 이질성, 다중 전하 스크린링 효과, 그리고 비정질 중간체의 동역학을 포함하면 실험과의 일치도가 크게 향상될 것으로 기대된다.
댓글 및 학술 토론
Loading comments...
의견 남기기