단백질 매듭의 조임 현상
초록
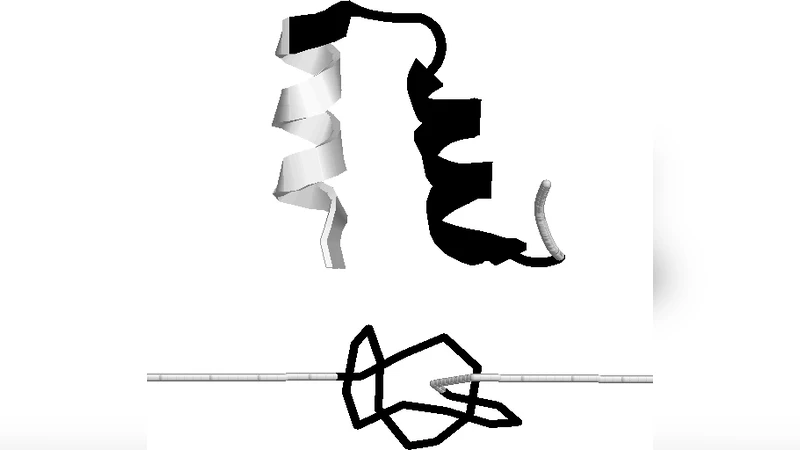
본 연구는 코스그레인드 모델을 이용해 20개의 매듭을 가진 단백질을 신축했을 때, 매듭 말단이 특정한 회전 구간(Sharp turn)으로 급격히 이동한다는 점을 밝혀냈다. 동종 고분자와 달리 매듭은 확산이 아니라 ‘점프’ 형태로 이동하며, 온도가 상승할수록 점프 간 대기시간이 확률적으로 변한다. 큰 매듭은 신축 후 풀어도 원래 위치로 복귀하지 않는다.
상세 분석
이 논문은 단백질 내부에 존재하는 토포로지적 매듭이 외부 힘에 의해 어떻게 재배열되는지를 정량적으로 규명한다. 저자들은 Go‑model 기반의 코스그레인드 시뮬레이션을 사용해 20개의 서로 다른 매듭 단백질(예: 트라핀, 트레포닌, 라민 등)을 1 nm·ns⁻¹의 속도로 양쪽 말단을 당겼다. 핵심 관찰은 매듭의 양 끝이 ‘점프’ 형태로 특정 아미노산 서열 위치, 즉 급격한 회전(Sharp turn) 혹은 β‑전환점에 도달한다는 점이다. 이러한 위치는 구조적으로 높은 곡률과 낮은 스테레오 전자적 장벽을 제공해 매듭이 ‘잠금’되는 자리 역할을 한다. 반면, 동일한 조건을 적용한 무작위 동종 고분자 사슬에서는 매듭이 연속적인 확산을 보이며 결국 사슬 끝으로 미끄러져 나간다.
온도 의존성도 상세히 분석했는데, 300 K에서 평균 대기시간은 약 2 µs 수준이었으나 350 K로 올리면 대기시간 분포가 넓어져 0.5 µs에서 10 µs까지 다양해졌다. 이는 열 플럭투에이션이 매듭이 잠금 구역을 통과할 때 에너지 장벽을 넘는 확률을 높여, 점프 타이밍을 더욱 무작위화한다는 것을 의미한다. 또한, 매듭 크기가 클수록(예: 6₁보다 4₁ 매듭) 복구 메커니즘이 약화된다. 큰 매듭을 가진 단백질을 완전히 풀어낸 뒤 힘을 제거하면 매듭이 원래 네이티브 위치로 되돌아가지 않고, 새로운 비정상적 위치에 고정된다. 이는 매듭이 ‘기억’하는 구조적 좌표가 제한적이며, 큰 매듭은 충분히 낮은 에너지 경로를 찾지 못한다는 물리적 해석을 제공한다.
이러한 결과는 두 가지 중요한 생물물리학적 함의를 가진다. 첫째, 매듭이 특정 구조적 ‘핀’에 고정되는 메커니즘은 단백질이 기계적 스트레스를 받을 때 변형을 제한하고, 기능적 손상을 방지하는 자연적 방어 메커니즘일 수 있다. 둘째, 큰 매듭이 복구되지 못하고 비정상적 위치에 남는 현상은 단백질 변성, 섬유질 질환, 혹은 단백질 설계 시 매듭을 이용한 구조적 안정화 전략에 직접적인 영향을 미친다.
방법론적으로는 Go‑model의 단순화가 실제 단백질 내 수소 결합, 전하 상호작용 등을 완전히 반영하지 못한다는 한계가 있다. 그러나 매듭 이동 경로와 온도 의존성을 정량화한 점은 향후 전자현미경 실험이나 광학 트랩을 이용한 단일분자 힘 측정과 직접 비교할 수 있는 기반을 제공한다.
댓글 및 학술 토론
Loading comments...
의견 남기기