바이러스 캡시드 구조 확장: 스카와 톰슨 문제의 새로운 연결
저자들은 전자들이 구면 위에서 쿠론 반발을 최소화하는 톰슨 문제에서 나타나는 “스카”(5‑7‑5 혹은 복합 패턴) 현상이 바이러스 캡시드 조립 과정의 중간 단계에서 발생할 수 있다고 가정한다. 스카가 형성될 때 축적된 굽힘 에너지가 스트레칭 에너지로 전환되면 icosadeltahedral 형태에서 원통형·원뿔형·비대칭 등 다양한 비구형 캡시드로 변형될 수 있다. 인공 단백질 케이지 실험을 통해 스카의 존재와 그 효과를 검증할 수 있다.
저자: Alfredo Iorio, Siddhartha Sen
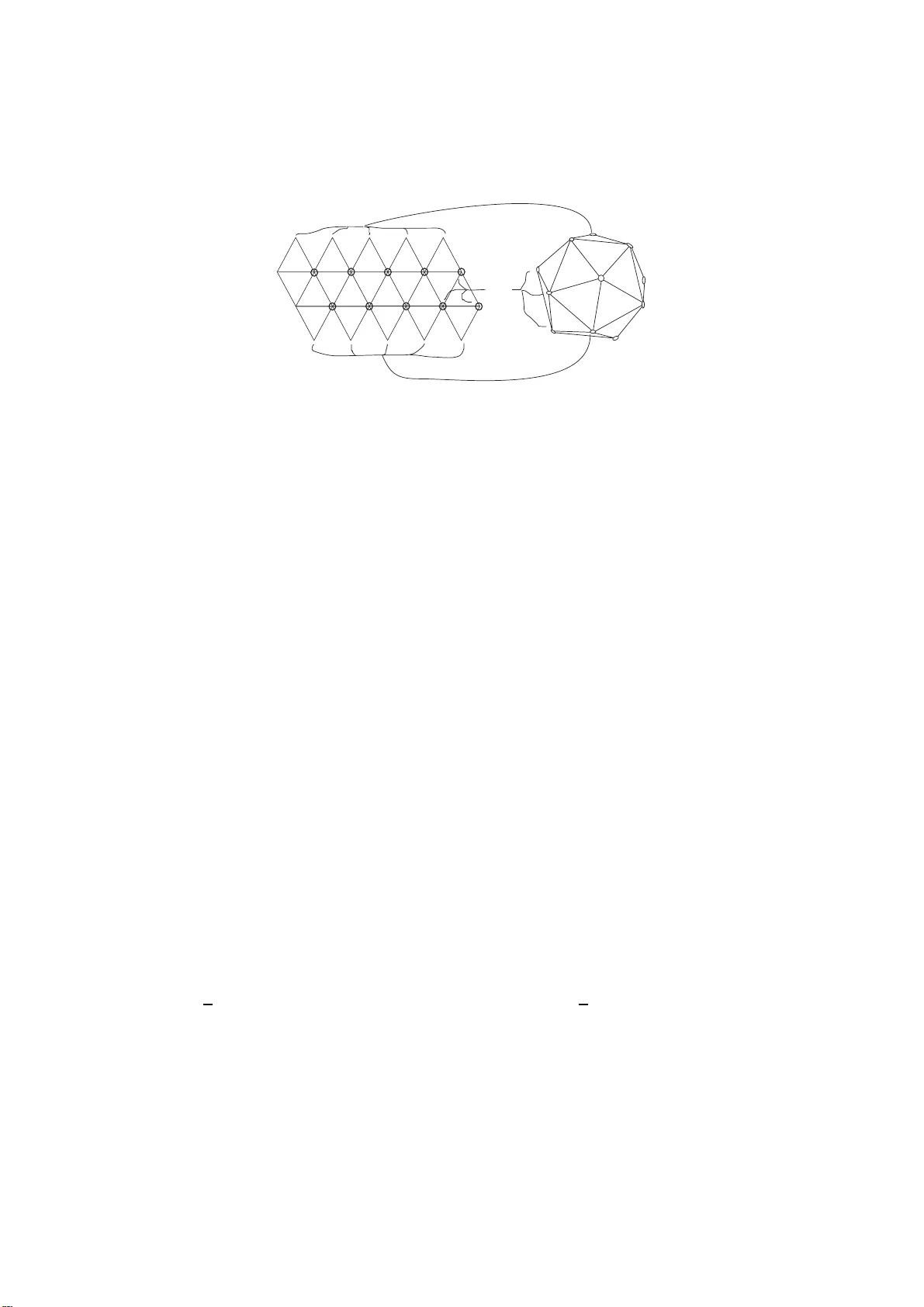
본 논문은 바이러스 캡시드 구조에 대한 기존의 Caspar‑Klug (CK) 모델을 물리학적 결함 이론과 결합해 확장하는 새로운 가설을 제시한다. 서론에서는 바이러스 캡시드가 유전물질을 보호하고 세포와의 결합에 중요한 역할을 한다는 점을 강조하고, 캡시드 형태를 크게 3가지(헬리컬, 이코사데랄, 복합)로 구분한다. 특히 이코사데랄 형태는 12개의 펜타머와 가변적인 헥사머로 이루어진 icosadeltahedron 구조이며, 이는 구면을 5‑각형과 6‑각형으로 타일링하는 토폴로지(오일러 식 χ=2)에서 도출된다. 저자는 CK 모델을 수학적으로 전개하여, (h,k) 정수쌍에 의해 정의되는 T=h²+hk+k²가 전체 단백질 수 N=60T을 결정하고, (h,k) 값이 캡시드의 ‘손성(chirality)’을 결정한다는 점을 설명한다.
다음으로, 전자들이 구면 위에 배치될 때 최소 전기적 반발 에너지를 찾는 고전적인 톰슨 문제를 소개한다. 톰슨 문제의 수치 해에서는 N=10T+2인 경우 12개의 오각형과 나머지 육각형이 규칙적인 삼각 격자를 이루지만, N이 충분히 클 때(대략 T≈10² 이상) 5‑6‑6 구조가 5‑7‑5 혹은 더 긴 5‑7‑5‑7‑5 형태의 “스카”(scar)로 변형되는 것이 에너지적으로 유리함을 보여준다. 스카는 한 쌍의 6‑각형을 5‑각형과 7‑각형으로 교체함으로써 국소적인 곡률을 +1/12와 –1/12로 바꾸지만, 전체 평균 곡률은 유지된다. 이러한 스카는 실험적으로도 폴리스티렌 구형 결정에서 관찰된 바 있다.
저자는 이 물리적 현상을 바이러스 캡시드 조립에 적용한다. 가설은 다음과 같다. (i) 초기 조립 단계에서 단백질이 CK 규칙에 따라 icosadeltahedral 형태를 만든다. (ii) 환경(세포 내 이온, 유전물질, 주변 단백질 등)으로부터 추가적인 굽힘 에너지(E_b)가 공급되면, 12개의 펜타머 중 하나 혹은 여러 개 근처에서 5‑6‑6 구조가 5‑7‑5 스카로 전환된다. (iii) 스카가 형성된 부위에서 굽힘 에너지가 스트레칭 에너지로 전환되면서 캡시드가 구형에서 원통형, 원뿔형, 혹은 비대칭 형태로 변형된다. 스카가 소멸하면 5‑7‑5가 다시 6‑6으로 돌아가지만, 이미 발생한 변형은 유지된다.
스카가 존재하는 경우와 없는 경우의 단백질 수는 동일하다(6+6=5+7), 다만 펜타머와 헥사머의 배치가 달라진다. 예를 들어, 하나의 단순 스카가 있으면 펜타머가 13개, 헵타머가 1개, 헥사머가 10T‑12개가 된다. 이러한 변형은 레트로바이러스와 같이 구형·원통형·원뿔형 캡시드를 모두 갖는 바이러스의 형태 변환을 설명하는 데 유용하다. 기존 연구(Ref.
원본 논문
고화질 논문을 불러오는 중입니다...
댓글 및 학술 토론
Loading comments...
의견 남기기