A Neurocomputational Account of Flexible Goal-directed Cognition and Consciousness: The Goal-Aligning Representation Internal Manipulation Theory (GARIM)
Goal-directed manipulation of representations is a key element of human flexible behaviour, while consciousness is often related to several aspects of higher-order cognition and human flexibility. Currently these two phenomena are only partially inte…
Authors: Giovanni Granato, Gianluca Baldassarre
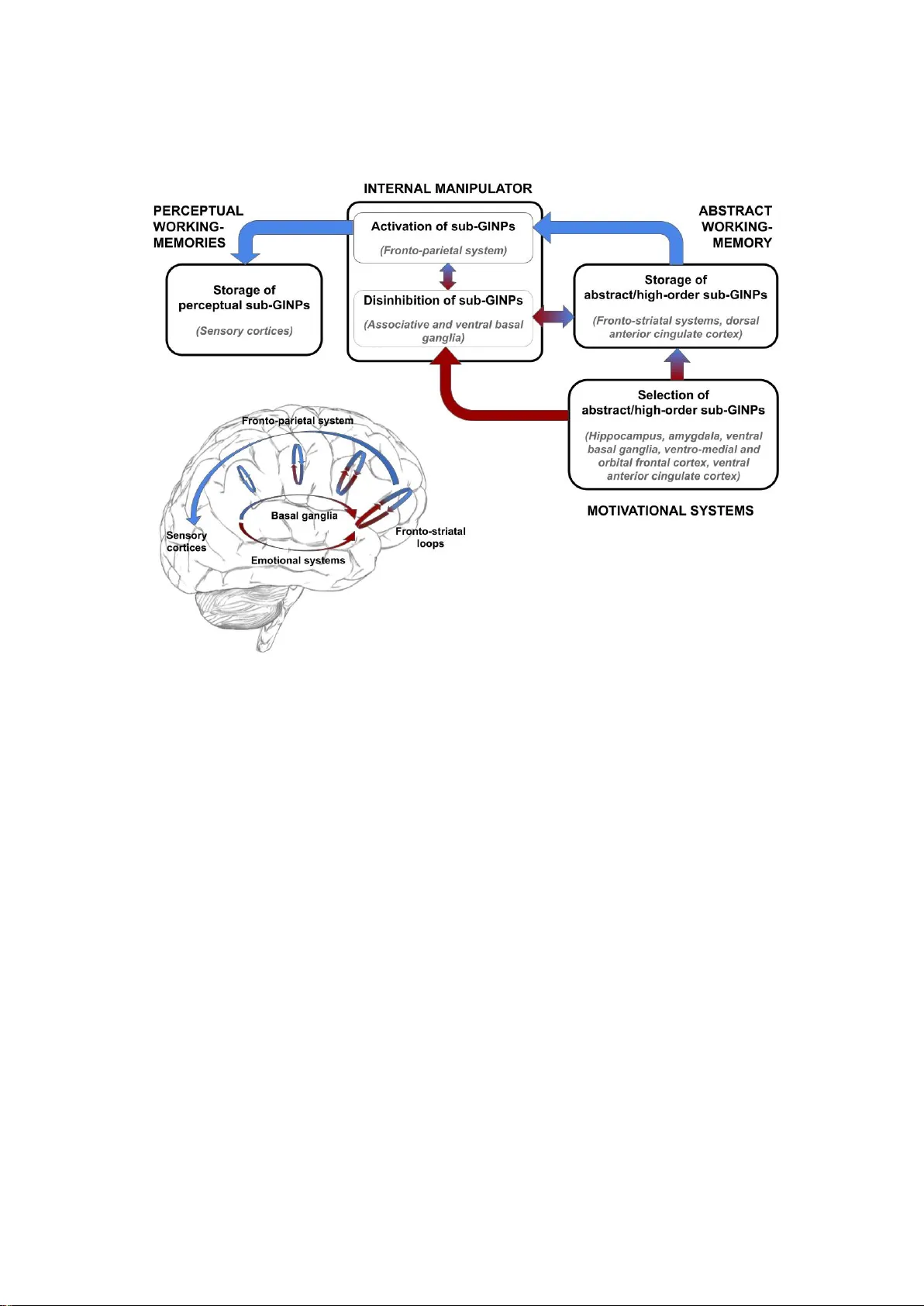
A Neuro computational Accoun t of Flexible Goal-directed Cognition and Consciousness: The Goal-Aligning Represen tation In ternal Manipulation Theory (GARIM) Gio v anni Granato a, ∗ , Gianluca Baldassarre a a L ab or atory of Emb o die d Natur al and A rtificial Intel ligenc e, Institute of Co gnitive Scienc es and T e chnolo gies, National R ese ar ch Council of Italy, R ome, Italy Abstract Goal-directed manipulation of represen tations is a key elemen t of human flexible behaviour, while con- sciousness is often related to several asp ects of higher-order cognition and h uman flexibilit y . Curren tly these t wo phenomena are only partially integrated (e.g., see Neurorepresen tationalism) and this (a) limits our understanding of neuro-computational pro cesses that lead conscious states to pro duce flexible goal- directed b eha viours, (b) preven ts a computational formalisation of conscious goal-directed manipulations of representations o ccurring in the brain, and (c) inhibits the exploitation of this knowledge for mo delling and technological purp oses. Addressing these issues, here we extend our ‘three-comp onen t theory of flex- ible cognition’ b y prop osing the ‘Goal-Aligning Representations In ternal Manipulation’ (GARIM) theory of conscious and flexible goal-directed cognition. The central idea of the theory is that conscious states supp ort the active manipulation of goal-relev an t internal representations (e.g., of w orld states, ob jects, and action sequences) to mak e them more aligned with the pursued goals. This leads to the generation of the kno wledge whic h is necessary to face nov el situations/goals, thus increasing the flexibilit y of goal-directed be- ha viours. The GARIM theory in tegrates k ey aspects of the main theories of consciousness into the functional neuro-computational framework of goal-directed b eha viour. Moreo ver, it tak es in to accoun t the sub jective sensation of agency that accompanies conscious goal-directed pro cesses (‘GARIM agency’). The prop osal has also implications for exp erimen tal studies on consciousness and clinical aspects of conscious goal-directed b eha viour. Finally , the GARIM theory can benefit tec hnological fields suc h as autonomous rob otics and mac hine learning (e.g., the manipulation process may describ e the op erations p erformed b y systems based on transformers). 1 The authors hav e equally contributed to the pap er. 1. In tro duction Goal-directed processes are at the basis of h uman flexible b eha viour. W e recently prop osed and v ali- dated the ‘three-comp onen t theory of flexible cog- nition’ (Granato et al., 2020; Granato and Baldas- sarre, 2021; Granato et al., 2022a; Granato et al.), ∗ I am corresponding author Email addresses: giovanni.granato@istc.cnr.it (Giov anni Granato), gianluca.baldassarre@istc.cnr.it (Gianluca Baldassarre) highligh ting that a goal-directed top-do wn manip- ulation of represen tations is at the basis of cogni- tiv e flexibility . Although the our theory success- fully describ es the neuro-cognitive pro cesses at the basis of cognitive flexibilit y , it does not focus on the higher-order pro cesses underpinning flexible cogni- tion (e.g., Planning and Problem Solving) nor on the role of consciousness in suc h pro cesses. Consciousness is a v astly debated concept and man y theories formalise its key features (Seth and Ba yne, 2022), which fo cus on different asp ects suc h as the integration of information (T ononi, 2008; T ononi et al., 2016; Koch et al., 2016; T ononi, 2004); the hierarchical conv ergence and div ergence zones elab orating cognitiv e/emotional brain information (Damasio, 1989; Meyer and Damasio, 2009; Dama- Pr eprint submitted to Neur al Networks Octob er 30, 2023 sio and Meyer, 2009a); the selection of relev ant in- formation into a central workspace and its ‘broad- casting’ to p eripheral areas (Baars, 1997; Baars et al., 2003; Baars, 2005; Baars et al., 2013); the top-do wn activ ation of multiple hierarchical brain systems b y the fron toparietal system (Dehaene et al., 1998a; Dehaene and Naccache, 2001; De- haene and Changeux, 2011); the difference b etw een first-order and higher-order representations (Brown et al., 2019; Cleeremans, 2011); the co ordination of effectiv e brain-b ody-environmen t sensorimotor in teractions (O’Regan and No e, 2001; O’Regan et al., 2005); the emergence of mult i-mo dal/mu lti- lev el representations that subserv e goal-directed b e- ha viours (Pennartz, 2015, 2018, 2022); the dynamic lo ops that generate and adjust predictions based on an inferential pro cess (Clark, 2013; Hohwy and Seth, 2020; F riston, 2018). These theories con- sider some k ey aspects that are commonly related to goal-directed cognition suc h as information hi- erarc hies, top-do wn information selection, sensory- motor interactions, multi-modal integration. Ho w- ev er, with the exception of neurorepresentational- ism (Pennartz, 2022), most theories capture only few elements of goal-directed cognition. This general p oor in tegration leads to several scien tific and technological issues. First, it limits the emergence of in tegrated frameworks that estab- lish the relationships b etw een consciousness, goal- directed cognition and flexible b eha viour. In par- ticular, it limits the understanding of the neuro- computational pro cesses that lead conscious states to pro duce a more flexible goal-directed b eha viour. Second, most theories of consciousness do not anal- yse the fine computational processes that supp ort a conscious and goal-directed manipulation of infor- mation. Although some theories ha ve led to com- putational mo dels of consciousness (e.g., Dehaene et al., 1998b; Pasquali et al., 2010; T ononi, 2008), there is not yet a clear description of the system- lev el manipulations of information that o ccur dur- ing conscious goal-directed pro cessing. Third, these limitations inhibit an effective and broad exploita- tion of these models and formalisations for tech- nological scop es. Although the emergence of in- terdisciplinary fields such as machine c onsciousness (Reggia, 2013) and consc iousness-inspired machine learning (e.g., Bengio, 2017), artificial in telligence (AI) and autonomous rob ot systems ha v e till re- cen tly sho wn rigid b eha viours and processes, in par- ticular ha ve failed to face no vel conditions or goals (Baldassarre and Granato, 2020; Hassabis et al., 2017; Lak e et al., 2017; Bubeck et al., 2023). Only recen tly some AI systems hav e exhibited examples of flexible general-purpose cognition (Li et al., 2022; Driess et al., 2023; Park et al., 2023). In terestingly , these mo dels rely on new algorithms (‘transform- ers’) that hav e notable links with the manipula- tion of internal represen tations. How ev er, the func- tioning of these systems is still p o orly understo od (Y un et al., 2021; Ab dou et al., 2021; Ag ¨ uera y Arcas, 2022; Sriv asta v a and al., 2022). Ov erall, these limitations negativ ely impact b oth scientific and technological adv ancements, which would b en- efit from a adequate integration betw een studies on goal-directed behaviour and higher-order aspects of consciousness. Addressing these issues, we extend here our ‘three-comp onen t theory of flexible cognition’ to higher-order aspects of goal-directed cognition and consciousness, thus introducing the Go al-A ligning R epr esentation Internal Manipulation (GARIM) theory of higher-order cognition and consciousness. The core idea of the GARIM theory is that con- scious pro cesses enhance the flexibility of goal- directed behaviour by supp orting the manipula- tion of goal-relev ant internal represen tations (e.g., of world states, ob jects, and action sequences). These manipulations generate the knowledge that the agen t lacks to improv e the alignment of such represen tations with the target goals, esp ecially when these are new or are pursued in nov el condi- tions. Therefore, a higher goal-oriented ‘alignmen t’ mak es activ e representations more likely to gener- ate successfully goal-oriented actions. The GARIM theory is based on five k ey features: (1) an adaptive function of consciousness; (2) sp e- cific representations at the basis of conscious and flexible goal-directed b eha viours (Goal-based In te- grated Neural P atterns; GINPs); (3) four in teract- ing neurofunctional systems supp orted by cortical net works and basal ganglia (hierarc hical perceptual w orking-memories, abstract working memory , in- ternal manipulator, motiv ational systems) at the basis of conscious goal-directed manipulations of GINPs; (4) four macro-classes of representation ma- nipulations which generate the goal-oriented miss- ing kno wledge (‘abstraction’, ‘specification’, ‘de- comp osition’, ‘comp osition’); (5) the emergence of a sub jective sensation of agency related to the rep- resen tation manipulations (‘GARIM agency’). This last feature contributes to differen tiate the concept of agency that emerges during a goal-directed b e- ha viour from those related to consciousness and 2 conscious states. Although the GARIM theory in- tro duces many considerations ab out low-order cog- nition (e.g., motiv ational and emotional asp ects of conscious flexible b ehaviour), it mainly focuses on higher-order asp ects of conscious cognition (e.g., top-do wn atten tion and planning). This is con- sisten t with the theory functionalist approach and tec hnological implications, as we sho w by linking it to concepts on higher-order human cognition and artificial intelligence (see section ‘GARIM theory and in telligence’). F or the same reason, here w e fo cus on meso-scale asp ects of the brain (e.g. in- teractions of brain macro-systems and broad neu- ral represen tations) rather then on neuron-level as- p ects of it. This level of analysis is suitable for ad- dressing the functions and computations targeted here, thus both for studying human cognition and for the developmen t of artificial in telligent systems and rob ots. The GARIM theory primarily aims to clarify the neuro-computational pro cesses that activ ely lead to more flexible goal-directed b ehaviours. In this resp ect, the theory can b e conceiv ed as a neur o- c omputational fr amework of c onscious and flexi- ble go al-dir e cte d c o gnition. Moreo ver, the theory giv es four sp ecific theoretical con tributions. First, it clarifies some asp ects of sub jectiv e exp erience and agency . In particular, it prop oses the concept of the GARIM agency to explain the different sub jec- tiv e exp eriences that accompany conscious states. Second, the theory con tributes to specify the neu- ro computational mechanisms underlying the main theories on consciousness. This allo ws the integra- tion of those mechanisms within a common func- tional and computational framework that pivots on goal-directed pro cesses. Third, the theory gener- ates insights for the exp erimental and clinical fields related to conscious states and goal-directed b e- ha viours. In particular, the theory is shown to b e compatible with relev ant exp erimen tal predic- tions of other theories on consciousness. Moreo ver, the theory gives indications for building new exper- imen tal paradigms for testing consciousness. Last, the theory offers an interpretation of the relation- ships betw een certain clinical impairmen ts and con- scious goal-directed b eha viours. F ourth, the the- ory provides some insights that could be useful for building new computational mo dels, ML-based sys- tems, and rob otic arc hitectures. Also, it can be use- ful to analyze existing ones. In this regard, compu- tational mo dels can op erationalise the theory , al- lo wing it to b e corrob orated with sp ecific empirical data and more detailed comparisons with other the- ories. On the other hand, indications to wards ML and rob otics could improv e the goal-directed flex- ibilit y of current systems and their interpretations (Baldassarre and Granato, 2020; Hassabis et al., 2017; Lake et al., 2017). Figure 1 summarises the main contributions of the GARIM theory and the organisation of this w ork. W e first describ e the three-component the- ory of flexible cognition, highlighting its key fea- tures, limitations, and also related technological fields (in particular AI and robotics). Building on suc h theory , w e then introduce the key features of the GARIM theory . Next, w e compare our prop osal with the ma jor theories of cons ciousness, sho wing that k ey higher-order asp ects of consciousness are captured by our integrated neuro-computational framew ork. W e then analyse the empirical implica- tions of the theory b y considering b oth exp erimen- tal and clinical evidence. Finally , we consider the implications for the design of new computational mo dels, AI systems and rob otic arc hitectures. Figure 1: The schema sho ws the k ey fields leading to the de- velopmen t of the GARIM theory and its main contributions. This schema also represents the ov erall organisation of this work. 2. The three-comp onen t theory of flexible cognition: k ey features, limitations and related fields The three-component theory of flexible cogni- tion formalises the neuro-cognitive pro cesses that b oost cognitive flexibility during goal-directed b e- ha viours (Granato and Baldassarre, 2021; Granato et al., 2020). In particular, the theory describ es b eha viours based on learnt action-outcome asso ci- ations and goals (Balleine and Ostlund, 2007; Thill et al., 2013; P assingham and Wise, 2012; Tsujimoto et al., 2011). In this context, ‘goals’ are defined as 3 represen tations of desirable future states that (a) can b e stored and internally re-activ ated and (b) can lead to select actions directed to achiev e them. Moreo ver, the theory op erationalises ‘cognitiv e flex- ibilit y’, an executiv e function that is defined as the capacit y of switc hing betw een different represen ta- tions dep ending on external and internal feedbac k (Diamond, 2013). The main idea of the three-comp onen t the- ory is that flexible c o gnition dep ends on the top- down go al-dir e cte d manipulation of r epr esentations . These manipulated representations b etter supp ort sensory–motor in teractions with the environmen t, thereb y bo osting cognitive flexibility and goal- directed b ehaviour. The theory proposes that three main elements are at the basis of flexible goal-directed b ehaviour (T able 1). Main elements Explanation Three k ey components Three neuro-functional systems support the goal-directed top-down manipulation of representations: · Executive W orking-Memory · Hierarchical perceptual systems · T op-down manipulator First-order and second-order representations/manipulations Two levels of representations and manipulations: · First-order (e.g., p erceptions and selective attention) · Second-order (e.g., abstract goals and inner-sp eech) Embodied sensory-motor loops Representation manipulations and sensory-motor embo died lo ops support goal-directed b eha viours T able 1: The table sho ws the main elements of the three- component theory of flexible goal-directed cognition. First, three neuro-functional systems support the goal-directed representation manipulation. An Ex- e cutive Working-Memory , supp orted by the pre- fron tal cortices (PF C) and frontal-striatal lo ops, stores the goals/sub-goals (Hartley and Sp eer, 2000; Bra ver and Bongiolatti, 2002). Multiple hier ar chi- c al p er c eptual systems , supp orted b y cortical p er- ceptual pathw ays, extract and store the goal-related represen tations (Rizzolatti and Matelli, 2003; Gaz- zaley and Nobre, 2012; Raffone et al., 2014a). A top-down manipulator , supp orted by frontal- parietal cortical system and basal ganglia-thalamo- cortical lo ops, applies a goal-directed manipulation of the stored representations at different stages of hierarc hical systems (V ossel et al., 2014; Parks and Madden, 2013; Redgrav e et al., 1999; Seger, 2008; Chelazzi et al., 2013; Pessoa, 2015). Note that fur- ther systems can act as a top-down manipulator (e.g., the inner-speech system; Granato et al., 2020, 2022a; Granato et al.). Second, the theory distinguishes b et w een ‘first- order’ and ‘second-order’ represen tations and ma- nipulations (Granato et al., 2020). The first term refers to perceptual representations and their ma- nipulations (e.g., visual selective atten tional pro- cesses). The second term refers to abstract/amo dal represen tations (e.g., goals/sub-goals, actions) and their manipulations (e.g., splitting of goals into sub- goals). Both refer to a self-directed form of manip- ulation at different levels of abstraction. Third, a synergistic interpla y betw een the goal- directed representation manipulation and embo d- ied sensory-motor lo ops is central to express flex- ible goal-directed b ehaviour. In particular, goal- directed b eha viour is supported by m ultiple manip- ulations of in ternal representations and the external w orld. The three-comp onent theory is v alidated trough an integrated theory-driv en/data-driven computa- tional approach. In particular, w e dev elop ed a neuro-inspired computational model based on this theory and we tested it with a neuropsychological test of cognitiv e flexibility (for further details see the section ‘T ow ards computational mo dels of the GARIM theory’). The computational mo del has re- pro duced the b ehavioural data obtained from v ari- ous cohorts of h uman participants, b oth in healthy and clinical conditions (Granato and Baldassarre, 2021; Granato et al., 2020, 2022a; Granato et al.). 2.1. Beyond the thr e e-c omp onent the ory Although the three-comp onen t theory has re- ceiv ed an exp erimen tal and computational v alida- tion, it shows limitations. First, although execu- tiv e functions and goal-directed b eha viours are of- ten linked to explicit cognition and consciousness, the three-comp onen t theory do es not take into ac- coun t the role of conscious processes in flexible cognition. Despite this limitation, the theory for- malises some key neuro cognitiv e pro cesses that are cen tral to the emergence of conscious states, such as information hierarchies (Damasio and Meyer, 2009b), top-down information selection (Dehaene 4 et al., 2011), sensory-motor in teractions (O’Regan et al., 2005), and first-order/second-order repre- sen tations (Brown et al., 2019). Second, the the- ory do es not formalise monitoring pro cesses and related higher-order goal-directed behaviours suc h as ‘planning’, defined as a flexible assem bling of new action sequences to accomplish goals (Pfeif- fer and F oster, 2013; Delatour and Gisquet-V errier, 2000), and ‘problem solving’, defined as a planning pro cess that inv olves partial knowledge (Newman et al., 2003). These limitations are due to the fact that the theory do es not tak e into consideration three key asp ects of higher-order cognition: w orld mo dels, ‘motiv ations’, and ‘emotions’. The follo w- ing paragraphs briefly articulate these concepts and their role in goal-directed b eha viour. World mo dels. The theory takes into accoun t the concept of ‘goals’, formalised as stored w orking- memory representations that c hange ov er time, but it do es not consider the concept of world mo del (Mars et al., 2011; Passingham and Wise, 2012; F uster and Bressler, 2015). W orld mo dels are rep- resen tations of the spatiotemp oral dynamics of the en vironment, integrating knowledge on the ev olu- tion of the physical en vironment and the effects that actions cause on it (Soltani and Koechlin, 2022). W orld mo dels supp ort planning and prob- lem solving as they allow the agent to internally sim ulate the dynamic transitions of the environ- men t, from a starting state to final goals. There- fore, b y in tegrating percept, goals and spatiotempo- ral simulations, they supp ort monitoring/ conflict resolution pro cesses and drive goal-directed plans (Huddy and Mansell, 2023; P ow ers, 2016). W orld mo del representations and processes are mostly supp orted by PFC systems and their lo ops with sub-cortical structures (in particular basal ganglia- thalam us and hipp ocampal systems; Houk et al., 1995; F uster, 2008; T ang et al., 2021; Patai and Spiers, 2021; Hasz and Redish, 2020). Motivations and emotions. Although the three- comp onen t theory implicitly assumes that motiv a- tional signals act as feedbac k to c hange the stored goals, it do es not explicitly examine the role of mo- tivations and emotions at the basis of flexible cogni- tion. Motiv ations supp ort the formation and reacti- v ation of goal represen tations during goal-directed b eha viours, guide learning, contribute to select b e- ha viors to p erform and energise them (Mars et al., 2011). Motiv ations can be divided in to extrin- sic/ph ysiological motiv ations (e.g., for safet y , wa- ter, and fo o d), so cial motiv ations (e.g., for belong- ing to a group), and intrinsic motiv ations (e.g., no velt y , surprise, competence impro vemen t, serving kno wledge and skills acquisition) (Panksepp, 1998; Gangestad and Greb e, 2017; Ryan and Deci, 2000; Mirolli et al., 2010). The role of emotions is still a debated topic (Scherer, 2005; Cabanac, 2002). Man y studies suggest that they promote the pro- duction of adaptive b eha viours (e.g., engagement, a voidance, and so cial comm unication; Panksepp, 1998; Ekman and Davidson, 1994; Damasio, 1998). In general, emotions predisp ose the b ody and brain to get into sp ecific adaptiv e ov erall mo des of func- tioning. A bulk of studies inv estigated neural corre- lates of motiv ations and emotions (P anksepp, 1998; Sc hultz, 2002; F rith, 2007; Amaral, 2002; Rolls, 2004; Lisman and Grace, 2005; Paus, 2001; O’Reilly et al., 2013; Ribas-F ernandes et al., 2011). These studies show that they are supp orted by many inter- acting sub-cortical structures (e.g., hypothalamus, am ygdala, insula, hipp o campus) and cortical struc- tures (e.g., medial/temp oral cortex, orbitofrontal cortex, an terior cingulate cortex, prefron tal cortex). Note that the terms ‘motiv ations’ and ‘emo- tions’ can refer to a global brain state (motiv ational state; e.g. sc hizophrenia patients sho w a low mo- tiv ational state defined ‘apath y’; Bortolon et al., 2018) or lo calised even ts (motiv ational signal; e.g., dopamine bursts supp ort information selection and learning; Berke, 2018). The t wo elements in ter- act (e.g., apath y in sc hizophrenic patients lead to an inefficien t information selection; Bortolon et al., 2018) and con tribute to shap e conscious states and goal-directed cognition. How ev er, in this w ork we particularly refers to the second function (motiv a- tional/emotional signals at the basis of the infor- mation selection comp onen ts). 2.1.1. Contributions of A rtificial Intel ligenc e and R ob otics AI and autonomous rob otics give important in- puts to extend the three-component theory to wards the GARIM theory . In particular, they b oth con- tribute to the in vestigation of brain and cogni- tion by supp orting their computational modelling (see section ‘T ow ards computational models of the GARIM theory’). A t the same time, as discussed in section ‘T ow ards AI systems and robotics architec- tures inspired by the GARIM theory’, they might b enefit from the scientific knowledge on brain and cognition to build more efficien t and flexible intelli- 5 gen t machines (Baldassarre et al., 2017; Baldassarre and Granato, 2020). A rtificial intel ligenc e, machine le arning, and neu- r al networks. Goal-oriented pro cesses hav e alwa ys pla yed a central role in artificial intelligence (Rus- sell and Norvig, 2016). In particular, AI has alwa ys attributed to human intelligence the primarily role of accomplishing goals through the search of the most suitable starting state-goal sequence (Simon, 1975). In terestingly , the t yp e of encoding of the rep- resen tations of planning elements strongly affected the evolution of the field. In particular, initial prob- lem solving systems used ‘atomic’ representations (i.e., distinct sym b ols for states and actions), which made the action sequence searc h inefficien t due to com binatorial explosion. Later, studies on planning (Russell and Norvig, 2016) ‘factorised’ the repre- sen tations of states into elements (e.g., ‘ob jects’) and relations b etw een elements (e.g., ‘b eing part of ’, ‘b eing on’). This change reduced the compu- tational costs at the basis of the action-sequence searc h. In parallel, ‘connectionist approac hes’ based on neural-netw orks proposed alternativ e systems based on ‘sub-sym b olic representations’, namely represen tations of features enco ded in neural pat- terns (McClelland and the PDPResearchGroup, 1986). Neural netw orks, initially used in machine learning to implement ‘reactiv e processes’, ha ve b een recently used to implement goal-directed pro- cesses such as planning (e.g., Rehder et al., 2018; W a yne et al., 2018). F urthermore, recen t research has prop osed that ‘deep neural net w orks’ could mo del k ey pro cesses underlying h uman conscious- ness (Bengio, 2017). Ov erall, artificial intelligence studies highligh t ho w higher-order cognition can b enefit of disentan- gled and factored representations that can (a) b e com bined in to new ones and (b) represent interde- p endencies among their sub-parts. Machine c onsciousness: key elements at the b asis of higher-or der c o gnition and c onsciousness. Ma- c hine consciousness (MC) is a research field aiming to define the key elements that artificial-in telligence and rob otic systems should hav e to exhibit a certain lev el of consciousness (Aleksander, 1995; Gamez, 2008). MC adopts b oth scientific and tec hnologi- cal approaches to accomplish this ob jective (Reg- gia, 2013). The scientific approach aims to de- v elop and v alidate computational mo dels built on the basis of the main theories of consciousness. The tec hnological approach aims to integrate el- emen ts of consciousness in to AI and rob otic sys- tems to improv e their flexibilit y and adaptability . Aleksander and Dunmall (2003) prop oses fives ‘ax- ioms’, stating which fundamental capabilities an in- telligen t system should hav e to exhibit a minimal lev el of consciousness:‘depiction’ (i.e. the capacity to represen t elements of the world), imagination, atten tion, planning, emotions. On the other hand, Gamez (2008) prop oses that MC systems can b e group ed into four classes based on their ‘conscious- ness simulation level’. A first class (MC1) inv olves the systems that exhibit a ‘conscious like’ exter- nal b ehaviour, such as AIs that exhibit h uman- lev el comp etence in playing complex games (F er- rucci, 2012; Lewis, 2012). A second class (MC2) encompasses systems that are generally inspired by theories of consciousness and sho w internal ‘cogni- tiv e pro cesses’ similar to those of conscious agen ts (e.g., atten tional pro cesses, motiv ation, world mod- els; Kugele and F ranklin, 2021; F ranklin et al., 2012; Holland, 2007; Marques and Holland, 2009; J¨ an tsch et al., 2010). The third class (MC3) in- v olves systems that are inspired by theories of con- sciousness and show brain-inspired arc hitectures (Dehaene et al., 2003; Gamez, 2010). The fourth and final class (MC4) encompasses systems able to engage in phenomenological forms of conscious sub jective exp erience. There is a hot debate re- garding the implementation of this kind of artificial systems (Carter et al., 2018; Reggia, 2013), but for no w no artificial system seems to b e able to un- dergo a human-lik e conscious internal exp erience. A t last, a relev ant review (Reggia, 2013) highlights that the prop osals of MC can b e categorised in five k ey classes, built on specific core principles: (1) in- ternal mo dels of the agen t itself (self-modelling); (2) information broadcasting; (3) higher-order rep- resen tations; (4) attention processes; (5) informa- tion integration. These frameworks (the fundamental axioms, the lev els of simulation, and the main implementa- tion principles) supp ort the formalisation of higher- order cognition and consciousness. All these ele- men ts hav e b een taken in to account to develop the GARIM theory . 6 3. The Goal-Aligning Represen tation Inter- nal Manipulation theory This section presen ts the fiv e ma jor elements of the GARIM theory (see T able 2): (a) the adaptiv e function that consciousness plays in goal- directed pro cesses; (b) sp ecific neural patterns that form conscious goal-related representations; (c) four anatomo-functional macro-systems that sup- p ort the manipulation of representations; (d) four classes of computational op erations that describ e suc h manipulations; (e) an explanation, based on the concept of GARIM agency , that links agency and sub jective conscious exp erience. The follo wing sections present these elements in detail. Main elements Explanation Adaptive function of consciousness Consciousness improves flexibility: conscious states and processes supp ort representation manipulations in order to increase their alignment with pursued goals, thereby enhancing goal-directed b eha viours. Goal-related representations Conscious goal-directed behaviours are supp orted by goal-directed integrated neural patterns (GINPs) having two key dimensions, i.e. goal-relevance and consciousness level. F our k ey components F our neuro-functional systems supp ort the goal- directed representation manipulation: abstract working-memory , perceptual working-memory , internal manipulator, motiv ational system. GARIM operations Goal-directed manipulations modify the GINPs and are sub jectively exp erienced as intentionally directed op erations. They are divided in four classes of operations: abstraction, sp ecification decomposition, comp osition. GARIM agency GARIM agency emerges during the execution of conscious flexible goal-directed represen- -tation manipulations. These generate a sub- -jective internal reality featured by three key elements: self-models, emotional and perceptual vividness, and manipulation control. T able 2: The five ma jor elemen ts of the GARIM theory . 3.1. The adaptive function of c onsciousness The GARIM theory p ostulates that the adap- tive function of c onsciousness is the impr ovement of flexibility during the expr ession of go al-dir e cte d b ehaviours. In p articular, c onscious states en- able agents to manipulate their internal r epr esen- tations (e.g., p er c eptions, thoughts and actions) in or der to gener ate know le dge mor e aligne d with the set go al; the higher go al-r elate d alignment le ads to mor e suc c essful go al-oriente d actions in famil- iar or novel situations, ther eby enhancing go al- dir e cte d b ehaviours (e.g., de cision-making, planning and pr oblem-solving). This feature is consistent with the commonly recognised role of goal-directed cognition for hu- man daily-life b ehaviours. Indeed, in case of new goals or situations habitual behaviours are often no longer suitable or efficien t. Moreo ver, also in case of b ehaviours that successfully lead to sp ecific sub-goals (e.g., an impro vemen t of physical p erfor- mance due to doping substances), they could show a misalignmen t with higher-level goals (e.g., honestly get an Olympics medal). Our proposal sp ecifies that represen tation manipulations generate more suitable kno wledge (e.g., plans, ob jects’ represen- tations, sub-goals) to face these conditions. Most theories of consciousness highlight that conscious pro cesses hav e an adaptive role for hu- man b eha viours (see section ‘Comparisons of the GARIM theory with other theories’). F or example, global w orkspace theories link conscious states to information sharing and amplification at the basis of decision making. On the other hand, Predic- tiv e Pro cessing theories link conscious processes to a con tinuous optimisation of inference/prediction mec hanisms related to goal-directed b eha viours. Imp ortan tly , the Neurorepresentationalism’s frame- w ork explicitly links Consciousness to goal-directed b eha viours. In particular, it suggests that con- scious processes generate the b est representations that serve goal-directed pro cesses. Our prop osal is compatible with studies on goal- directed b ehaviour and consciousness. Ho wev er, it prop oses a sp ecific fo cus on higher-order pro cesses. F or example, Neurorepresentationalism fo cuses on the emergence of represen tations that are subse- quen tly exploited b y goal-directed pro cesses. In- stead, the GARIM theory formalises mechanisms that op erate and constitute goal-directed cognition (i.e., goal-directed representation manipulations). 3.2. Go al-b ase d Inte gr ate d Neur al Patterns (GINPs): c onscious r epr esentations at the b asis of flexible go al-dir e cte d b ehaviour The GARIM theory describ es brain states that supp ort higher-order conscious cognition, leading to the expression of flexible goal-directed b eha viour. T o adequately represent the p eculiarities of these 7 states, we introduce the concept of ‘Go al-b ase d In- te gr ate d Neur al Pattern ’ (GINP; see Figure 2). Al- though we cannot present direct exp erimen tal evi- dence for GINPs, their existence is compatible with studies on goal-directed b eha viour and other theo- ries of consciousness (see the paragraph ‘GINPs and other definitions of conscious representations’). Definition, fe atur es and br ain c orr elates. W e define a GINP as an active distributed neural represen- tation that is characterised by tw o features: (a) c onsciousness level : it is consciously p erceiv ed and th us in tentionally manipulable; (b) go al-r elevanc e : it is functionally relev ant for the pursued goals. GINPs are integrated representations that hav e a comp ound nature, in particular are formed by sub- parts (‘sub-GINPs’). These sub-GINPs enco de dif- feren t asp ects of goal-directed conten ts (e.g., p er- cepts, affordances, actions, goals). W e hypothesise that GINPs are encoded at mul- tiple levels b y man y structures in the brain hier- arc hies (see figure 2). A GINP related to a sp e- cific goal (e.g., ‘patting a dog’, enco ded in the PF C) could b e formed by sub-GINPs related to visual appearance (e.g., the dog asp ect, enco ded b y visual areas), an o vert sound (e.g., the bark- ing, enco ded by auditiv e areas), a inner-sp eec h pro duction (e.g., the word ‘dog’, enco ded by lan- guage areas), and p ossible related actions (e.g., ‘patting’, enco ded b y motor areas). W e expect that the strength of ph ysical/functional connections b e- t ween sub-GINPs (i.e., neural integration) v aries dep ending on their consciousness lev el and goal- relev ance. Dynamically , only one GINP can become con- scious at a certain momen t. This is c onsisten t with the commonly accepted fact that only a limited represen tation can access consciousness at a time. Ho wev er, the GINP contin ually evolv es under the effect that representation manipulations hav e on its sub-GINPs. The integration b et ween the sub- GINPs could b e supp orted by b oth physical connec- tions and functional connectivity . Sub-GINPs can ha ve such a low level of in tegration that they stop forming in tegrated represen tations. In this case, for simplicit y , w e still k eep the ‘-GINP’ word ending (see ‘Non-GINPs’ in the next paragraph). Non-GINPs, Pr e-GINPs, T emp-GINPs and GINPs: fr om unc onscious to stably c onscious r epr esentations, and vic e versa. W e h yp othesise that GINPs enable representation manipulations, and th us conscious and flexible goal-directed b eha viours. How ever, on the basis of their con- sciousness level and goal-relev ance, w e define four kinds of representations that can emerge under the op eration of consciousness processes. GINPs : whole conscious representations that ha ve a high level of goal-relev ance and stability in time, and th us strongly affect goal-directed b e- ha viour. T emp-GINPs : representations that hav e a lo w goal-relev ance, but nevertheless temp orar- ily access consciousness (e.g., salien t stimuli such as unexp ected skin pressure or distracting internal though ts). They can acquire goal-relev ance, thus b ecoming GINPs, or can b e suppressed by top- do wn atten tion. Pr e-GINPs : unconscious repre- sen tations that hav e a high level of goal-relev ance but do not hav e the supp ort of top-down attention, th us remaining unconscious. They can be activ ated b y background pro cesses (e.g., priming) and can in- directly influence conscious represen tations. Non- GINPs : unconscious representations that hav e lit- tle or no goal-relev ance, but are activ ated b y ex- ternal even ts or related in ternal active represen- tations. Dep ending on their features, Non-GINPs could hav e a v ery lo w in tegration. Therefore, they could stop b eing ‘integrated global representations’ and b ecoming ‘scattered local representations’. In our descriptions, we will generally refer to the whole brain representations (GINPs) but their sub-parts (sub-GINPs) will often inherit their consciousness-lev el/goal-relev ance prop erties. F or example, when w e refer to a GINP as a ‘goal- relev an t consciously perceived representation’, we also imply its sp ecific sub-GINPs are goal-relev an t and accessible by consciousness pro cesses. Instead, when we consider the different status of the cur- ren tly active sub-GINPs, we refer to them sepa- rately . Figure 2 (box C) sho ws that the four t yp es of rep- resen tations can exhibit a sequential dynamic rela- tionship. F or example, a non-GINP could progres- siv ely acquire goal-relev ance until b ecomes a pre- GINP and then a GINP . Conv ersely , a GINP can b e de-activ ated and b ecome a pre-GINP; and in case it loses most of its goal-relev ance it b ecomes a non-GINP . On the other hand, a non-GINP can temp orally access consciousness with a low goal- relatedness, b ecoming a temp-GINP (e.g., repre- sen ting an ob ject that suddenly enters the field of view). How ev er, unless it is later recognised as rel- ev an t for the set goal, th us b ecoming a GINP , it is discarded and b ecomes a non-GINP . 8 Figure 2: (A) Different kinds of Goal-based Integrated Neural P atterns (GINPs), classified on the basis of their k ey dimensions: goal-relev ance and conscious level. (B) Possible neural correlates of a GINP; the figure shows four sub-GINPs (orange, grey , violet, yello w), enco ding different goal-relev an t elements (e.g., p erceptual features of ob jects, affordances, actions, abstract representations of goals). (C) P ossible sequen tial relationship b et ween differen t kinds of GINPs; green arrows indicate an ‘unconscious-to-conscious’ c hange (e.g., a pre-GINP that b ecomes a GINP) while red arrows indicate the in verse change. Note that a curved arrow, on top of each b ox, indicates the level of goal-relatedness/goal-based representation manipulation (e.g., absent, temp orary , prolonged). The difference b et ween non-GINPs/pre-GINPs and temp-GINPs/GINPs accounts for the differ- ences betw een subliminal/implicit/unconscious and supraliminal/explicit/conscious represen tations highligh ted by many brain studies (Meneguzzo et al., 2014). In addition, and imp ortan tly , these concepts can highlight the difference b et w een awar eness and c onsciousness . Indeed, non-GINPs and pre-GINPs can temp orarily access conscious- ness, th us b ecoming temp-GINPs or stable GINPs that the agen t can rep ort ab out (a wareness). On the other hand, only GINPs can b e the target of activ e manipulation op erations that, thereby supp orting higher-order consciousness and flexible goal-directed b ehaviour. Therefore, this distinc- tion implies that aw areness plays a ‘preparatory’ role for conscious goal-directed pro cesses, while consciousness inv olv es the core op erativ e stage of conscious goal-directed pro cesses. In section ‘A GARIM agency scale’ we refer to these different concepts as ‘phenomenal consciousness’ (aw are- ness), ‘access consciousness’, and ‘manipulative consciousness’. The scale implies that Awareness and Consciousness are t wo p oles of conscious goal-directed cognition. GINPs and other definitions of c onscious r epr e- sentations. Many theories of consciousness exp ect that only sp ecific represen tations hav e the proper- ties needed to b e consciously pro cessed (see section ‘Comparisons of the GARIM theory with other the- ories’ for more detailed comparisons). F or exam- ple, the IIT prop oses that conscious states show a high level of integration and sp ecification. The GWT/GNWT prop ose that consciousness is sup- p orted b y stable activ ations of highly integrated systems, dispatc hing information to short-range sub-mo dules. The Radical Plasticity Theory (part 9 of the Higher-Order Theories of consciousness) sug- gests that only the meta-represen tations, featuring stabilit y , strength and distinctiveness, are p erceiv ed as conscious conten ts. Finally , Neurorepresenta- tionalism p ostulates that conscious states dep end on multimodal/multi-lev el representations subserv- ing goal-directed b eha viours. While the GARIM theory fo cuses on a subset of conscious representations (i.e., those that sup- p ort goal-directed b eha viours), the definitions re- lated to the different types of GINPs is compati- ble with those prop osed by several theories of con- sciousness. In addition, ho wev er, the definition of GINPs explicitly requires their activ e goal-oriented manipulation to b ecome stably conscious. 3.3. F our key c omp onents at the b asis of go al- dir e cte d manipulations of r epr esentations The GARIM theory p ostulates that conscious higher-order cognition is supp orted by goal-oriented manipulations of representations. These manip- ulations rely on four key ‘comp onen ts’ (Figure 3), namely four partially ov erlapping anatomo- functional brain macro systems. Note that the GARIM theory inherits some key comp onen ts from its precursor theory , extending them (in particular, b y adding a fourth motiv ational comp onen t) and further specifying their functioning (e.g., see the section ‘The four classes of GARIM computational op erations’). (1) Per c eptual working memory c omp onent. The three-comp onen t theory proposes that p erceptual hierarc hies play a key role in goal-directed b e- ha viours. Moreov er, v arious theories of conscious- ness p ostulate that hierarchical perceptual sys- tems and p erceptual working-memories supp ort the emergence of conscious states (see section ‘Compar- isons of the GARIM theory with other theories’). In the GARIM theory , the p er c eptual working memory c omp onent pla ys a key role for the emer- gence of GINPs. The comp onent corresp onds to partially segregated ‘unimo dal’ sub-systems that p erform b ottom-up sensory processing. These op- erations support the formation of increasingly ab- stract p erceptual sub-GINPs (e.g., from lo w-level features to high-lev el representations). In par- ticular, the bottom-up information flows conv ey pre-GINPs/non-GINPs representations to higher- lev el cognitiv e areas. The comp onen t also sup- p orts a top-down information flo w, causing the re- activ ation of the peripheral sub-GINPs (e.g., goal imagination and mental simulations during visual planning). These manipulation pro cesses can con- tribute to transform pre-GINPs into GINPs and to inhibit temp-GINPs (non-GINPs that temporary access consciousness). The comp onent also imple- men ts p eripheral mo dal working memories. These main tain active p erceptual representations ha ving a short duration and a high lev el of detail (e.g., the p erceptual representation of a goal). In the brain, the comp onen t is supp orted by cor- tical hierarchical pathw ays. These enco de bottom- up information at m ultiple lev els of abstraction, in- stan tiating extensive asso ciative netw orks linking sub-GINPs enco ded in differen t cortices. At the same time, fronto-parietal cortical pathw ays acti- v ate in a top-down goal-directed fashion the con- ten ts of the mo dal working memories. (2) Abstr act working memory c omp onent. The three-comp onen t theory proposes that an abstract w orking memory plays a k ey role in goal-directed b eha viours (e.g., storage of abstract goals). More- o ver, most theories of consciousness ascrib e a cen- tral role to w orking memory (see section ‘Compar- isons of the GARIM theory with other theories’). In the GARIM theory , the abstr act working- memory c omp onent supports the activ e mainte- nance and integration of different goal-relev an t sub-GINPs (e.g., related to contexts, b ehavioural strategies, predictions, and v alues). These sub- GINPs are related to lo w-lev el sub-GINPs (e.g., b e- ha vioural strategies can b e related to mo vemen t represen tations and exp ected somatosensory and visual feedbac k) but encode more abstract informa- tion with respect to p erceptual sub-GINPs. This feature makes them a form of meta-kno wledge. Imp ortan tly , abstract sub-GINPs dynamically in tegrate both spatiotemp oral relations of the w orld elemen ts (e.g., o wn b ody parts, ob jects, other agen ts) and agen t’s predictions (e.g., goal-related action outcomes) in world mo dels . Thus, the ab- stract working-memory comp onen t exploits w orld mo dels to monitor plans (i.e., c heck the predic- tion correctness based on p ercept and goals/sub- goals), to counter in ternal (self ) and external (en- vironmen t) disturbances, and finally to generate the missing kno wledge (e.g., new p oin ts of view on ob- jects, new solutions). Within the brain, abstract multimodal sub- GINPs are enco ded b y differen t prefron tal cortices (e.g., dorsolateral PFC, v entrolateral PF C, and an- terior cingulate cortex) and related sub cortical ar- 10 Figure 3: Sc hema sho wing the ‘comp onen ts’ of the GARIM theory , and their relation with brain anatomo-functional systems. The red-to-blue coloured gradient indicates the decreasing involv ement of motiv ational/emotional pro cesses. eas (e.g., basal ganglia-thalamo-cortical lo ops and hipp ocampal system). Within eac h cortical area, neural winner-tak e-all mechanisms allow the acti- v ation of only one or few possible patterns at a time. Imp ortan tly , the abstract working memory comp onen t plays a ‘h ub role’ by putting in rela- tion sub-GINPs in different areas (e.g., different re- gions of the fronto-parietal netw ork). In particu- lar, it dynamically in tegrates abstract sub-GINPs with p erceptual sub-GINPs, thus realising a close in teraction with p erceptual working memory . This coupling supp orts p erceptual monitoring underly- ing conflict resolution, goal-alignmen t (e.g., percep- tual monitoring of goal-prediction matc hing), and sub-GINP sequences activ ations at the basis of vi- sual planning (images of world states tra versed to reac h the goal). (3) Internal manipulator c omp onent. The three- comp onen t theory prop oses that goal-directed b e- ha viours are supp orted by a top-down manipulator of representations. Several theories of consciousness attribute a central imp ortance to attentional pro- cesses and their top-do wn influence on conscious in- formation (see section ‘Comparisons of the GARIM theory with other theories’). The GARIM theory proposes that an internal manipulator c omp onent manipulates the con tents of abstract and p erceptual w orking-memories. In particular, it selects and warps p erceptual and ab- stract sub-GINPs to generate sequences of GINPs with increasing goal-alignment. Imp ortantly , these manipulations support (a) monitoring and align- men t of goals, sub-goals and world mo dels (ab- stract sub-GINPs) with p erceptions (p erceptual sub-GINPs) to solve conflicts and internal/external disturbance and (b) the subsequent generation of new kno wledge needed in case of nov el situations and goals. In the brain, the manipulator’s op erations are 11 supp orted b y t w o ma jor selection mec hanisms. One corresp onds to local inhibitory circuits of cortex, in particular those comp osing the cortical fron to- parietal system. The second corresp onds to the disinhibition mechanisms of basal ganglia-thalamo- cortical lo ops. The influence of basal ganglia on the cortex has a diminishing gradient, moving from fron tal to p osterior cortical areas. Although the GARIM theory fo cuses on these manipulation brain systems, others could con tribute to goal-directed manipulations. F or example, the three-comp onen t theory prop oses that the language system acts as an in ternal manipulator of abstract goals (e.g., in- ner sp eec h; Granato et al., 2020, 2022a; Granato et al.). (4) Motivational c omp onent. Motiv ational systems pla y a key role in the expression of goal-directed b eha viours, from goal formation to action selection (see section ‘Beyond the three-comp onen t theory’). V arious theories of consciousness tak e into consid- eration the role of motiv ational and emotional pro- cesses for consciousness (see section ‘Comparisons of the GARIM theory with other theories’). In particular, the GARIM theory proposes that a motivational c omp onent indirectly guides the ma- nipulator, contributing to select goals at differen t lev els of abstraction within the abstract working memory . Moreov er, the motiv ational component also directly contribute to the manipulator op er- ations, giving different salience to p erceptual and abstract sub-GINPs (see Figure 3). T o this pur- p ose, the motiv ational component closely interacts with the p erceptual and abstract working memories to p erform goal-monitoring and goal-aligning op er- ations based on the manipulation of p ercepts, w orld mo dels and plans. The motiv ational comp onen t also contributes to giving an emotional sub jective nuance to conscious represen tations. In particular, p erceptual sub- GINPs (e.g., representations of external stimuli and an ticipated outcomes) are ev aluated (appraisal) on the basis of their contribution to the achiev emen t of goals (goal-alignment). This pro cess contributes to in tegrate cognitiv e and emotional asp ects of goal- directed behaviour, and plays a key role for the agen t’s sub jective experience accompan ying con- sciousness (see section ‘GARIM agency and the sub jective exp erience of consciousness’). In the brain, motiv ational and emotional ev alua- tions drive the selection pro cesses of basal ganglia and cortical winner-take-all mec hanisms. In partic- ular, ev aluations generated in sub cortical structures (e.g., the h yp othalam us, amygdala, hipp ocampus) reac h the basal ganglia starting from the the limbic lo op. Moreov er, they reach v arious cortical areas starting from the PF C v entral areas (e.g., orbital, medial, and insular cortex). 3.4. F our classes of GARIM op er ations The interaction of the four comp onen ts supp orts goal-directed manipulations of internal representa- tions. These manipulations are divided in four GARIM op er ations (Figure 4). These op erations mo dify the GINPs and are sub jectively experienced b y the agen t as inten tionally directed operations (see section ‘GARIM agency and the sub jective ex- p erience of consciousness’). The four classes are no w considered in detail. Figure 4: The four classes of GARIM op erations that the manipulator performs on GINPs. (1) Abstr action. Abstraction causes the generation of sub-GINPs at different levels of abstraction, from p erceptual sub-GINPs to abstract sub-GINPs. Ab- straction also executes goal-dep enden t dimensional reductions, preserving only goal-relev an t asp ects of lo w-level sub-GINPs. F or example, in addressing the goal ‘grasping the cup’, abstraction op erations migh t change the detailed sub-GINP related to the p erceptual representation of the cup in to a more ab- stract goal-orien ted sub-GINP (e.g., a shape-based represen tation, ignoring colour b ecause it is not use- ful for the pursued goal). In the brain, abstraction relies on the hierarchi- cally organised stages of cortical path wa ys. Basal ganglia-thalamo-cortical macro lo ops (limbic, asso- ciativ e, motor) operate the selection of patterns at suitable levels of abstraction. 12 (2) Sp e cific ation. Sp ecification p erforms the in- v erse op erations with resp ect to abstraction. F or example, starting from an abstract sub-GINP (e.g., ‘something to drink with’) it can generate a sub- GINP corresp onding to a sp ecific ob ject (e.g., ‘m y preferred tea cup’). Since sp ecification inv olves mappings from a few to many features, it requires a goal-directed and con textualised generation of suitable information (e.g., the p erceptual details of ‘my preferred cup’ when the goal is ‘drink tea at home’). These op era- tions are made p ossible b y the manipulator’s selec- tions and by the generativ e netw orks of p erceptual and abstract working memories. In the brain, sp ecification relies on the top- do wn ‘in verse’ activ ation of cortical path wa ys, mo v- ing from m ultimo dal representations in the fron tal cortices to mo dalit y-sp ecific representations in the lo wer sensory cortices. The generation of more de- tailed representations is guided b y the cortical and basal-ganglia selection pro cesses. (3) De c omp osition. Decomp osition p erforms the separation of representations (GINPs and sub- GINPs) into sub-parts. This op eration executes a differen t kind of manipulation with resp ect to ab- straction and sp ecification. While the latter p er- form a ‘vertical manipulation’ that changes the ab- straction lev el, decomp osition p erforms a ‘horizon- tal manipulation’ at a fixed lev el of abstraction. F or example, decomp osition could extract the represen- tation of an ob ject (e.g., ‘a tea cup’) from the back- ground, or the representation of a part of the ob ject (e.g., ‘the handle’) from other parts (e.g., ‘the cup con tainer’). In the brain, decomp osition could b e supp orted b y neural structures similar to those of sp ecifica- tion, thus inv olving the cortex and basal ganglia- thalamo-cortical lo ops. How ev er, it might more strongly in volv e the channels and sub-channels within those lo ops to disinhibit specific cortical con- ten ts. Cortical lo cal winner-tak e-all mechanisms should facilitate the selection of sub-parts of neural patterns. (4) Comp osition. Comp osition p erforms the in- v erse op erations with resp ect to decomp osition, in- tegrating many sub-GINPs into larger sub-GINPs or into a coheren t whole GINP . Through comp osi- tion, the agent can build global items starting from its parts (e.g., to consider a ‘cup container’, ‘han- dle’, ‘tea’, and ‘tea sp oon’, as a whole ‘tea cup’). Comp osition supp orts v arious asp ects of goal- directed pro cesses. F or example, it supports the generation of plans (e.g., b y ch unking a sequence of actions and their effects) or imaginary pro cesses leading to solv e a problem (e.g., building a new to ol b y aggregating v arious parts). Comp osition p erforms a different manipulation with resp ect to abstraction. Abstraction performs a dimensional reduction (loss of information) while comp osition ‘c hunks representations’ at the same level of ab- straction. Ho wev er, comp osition and abstraction could give rise to adaptive synergies. F or exam- ple, they could lead to in tegrating many sub-GINPs at the same abstraction lev el, then transforming the resulting sub-GINP into a more abstract one (e.g., ch unking ‘reaching’, ‘grasping’, ‘transport- ing’, ‘drinking’ to generate the abstract goal ‘taking a tea’). In the brain, composition might rely on func- tional connectivity betw een different netw orks. Moreo ver, it might rely on physical connectivit y linking semantically related neural patterns (e.g., t wo differen t colours within the visual cortex, or the ‘red’ colour in the visual cortex and ‘alertness’ in an affective area). The inte gr ate d functioning of the GARIM op er a- tions: r epr esentation manipulations b o ost flexibil- ity during pr oblem solving. The GARIM op erations giv e rise to a sup er-ordinate function we call Con- scious Know le dge T r ansfer (CKT) . CKT refers to a transfer of knowledge from familiar contexts to no vel con texts, thus supporting flexible h uman cog- nition and b ehaviour. In particular, CKT op er- ates by flexibly abstracting, specifying, decomp os- ing, and comp osing the sub-GINPs that enco de the curren t knowledge (e.g., related to ob jects, goals, actions, and expected outcomes). Therefore, on the basis of m ulti-level goal-monitoring/goal-alignment ev aluations, CKT allo ws the agent to generate the necessary knowledge to improv e p erformance, to successfully act in changed conditions, or to ac- complish nov el goals. Differently from the concept of gener alisation , CKT leads to the generation of new knowledge b eyond previous experiences. While generalisation inv olves in terp olation pro cesses (e.g., the imagination of a goal p osition that in volv es an ob ject p ositioned b et ween tw o previously exp eri- enced p ositions), CKT inv olves extrapolation pro- cesses (e.g., the imagination of an ob ject lo cated an ywhere in a known space; or the generation of a new to ol based on comp osing elements). These op- 13 erations are based on the extraction of relev an t reg- ularities from previous experiences, and their trans- formation to generate knowledge to address nov el c hallenges (decisions, plans, problems). Problem solving tasks are b est suited to illus- trate the CKT and the GARIM operations. Such problems are challenging b ecause their solution re- quires the generation of missing knowledge on ill de- fined comp onen ts. F or instance, consider the clas- sic Dunck er’s problem (Dunck er, 1945). In this task participan ts are required to fix a candle on a wall. They can only use some pins, a v ailable in a b o x, and some matc hes to solve the problem. The solution requires to pin the cardb oard b ox on the w all and then set the candle on it. This solution requires a ‘c hange of p ersp ectiv e’ on the elements of the prob- lem (Guilford, 1967; Chrysikou et al., 2016). In- deed, participants generally consider the b ox only as a container, but this change of p ersp ectiv e leads them to focus on its properties (e.g., ‘cardb oard can b e pinned’). Thus, they discov er that the b o x can serv e as a candle holder. As highlighted by the ‘rep- resen tational change theory’ (Ohlsson, 1992), the solution requires the participant to generate a new suitable representation of the k ey problem’s sub- comp onen ts (e.g., of the b o x). The GARIM theory can explain the manipula- tion and generation of kno wledge that leads to the solution of the Dunck er’s problem. F or example, an agen t could use decomp osition to parse the scene, and then sequentially activ ate the sub-GINPs that enco de the different ob jects of the task. When fo- cusing on the cardb oard, the agent might use de- comp osition and sp ecification to analyse the dif- feren t feature-based sub-GINPs of the cardb oard (e.g., the usual function, the shape, and the mate- rial). These sub-GINPs, p otentially influenced by a con text-dep enden t priming effect (e.g., a pre-GINP enco ding the pin), can recall the represen tation of a previous exp erience (e.g., the agent that used pins to stic k cardb oard dra wings on the wall). Ex- ploiting comp osition, the agent might then trans- fer the piece of knowledge ‘cardb oard things can b e pinned on w alls’ (a sub-GINP) to the cardb oard b o x (another sub-GINP). A t last, the resulting sub- GINP could b e abstracted (abstraction) and com- pared with the initial goal of ‘attaching the can- dle to the wall’. A high corresp ondence b et ween the t wo would imply a high goal-alignmen t of the GINP , achiev ed thanks to the CKT. 3.5. Subje ctive exp erienc es during c onscious go al- dir e cte d b ehaviours: the GARIM agency The nature of sub jectiv e exp eriences is widely debated in the literature, whic h commonly refers to them as the ‘hard problem of consciousness’ (Chalmers, 1995). Although the GARIM theory do es not offer a solution to the hard problem of con- sciousness, it prop oses its own p erspective on this topic. In particular, the theory relates the activity of the in ternal manipulator with the emergence of a sub jective experience of agency . Therefore, we in- tro duce the concept of GARIM agency to identify the sense of agency that emerges during the expres- sion of conscious flexible goal-directed b eha viours. In particular, the theory prop oses that the ma- nipulation of representations generates an internal simulate d r e ality ha ving three k ey features: self- mo dels, emotional/perceptual vividness, and ma- nipulation control. First, the simulated realit y inv olves some aspects of the agent itself. This self-simulation can b e en- hanced based on previous exp eriences with other in tentional agen ts (F ernandez-Duque et al., 2000). Second, the manipulator activ ates low-lev el sub- GINPs that enrich the GINPs with detailed p er- ceptual representations. The GINPs are contin u- ously ev aluated with respect to their goal-related alignmen t and thus they are emotionally c harged. These GINPs hence exhibit p erceptual and emo- tional features similar to those that the agent ex- p eriences when acting in the environmen t. F or this reason, the internally sim ulated and manipulated realit y is vividly p erceiv ed and felt similarly to the real experience. Third, the in tentional manipula- tion of representations cause imagined effects simi- lar to those caused by motor actions p erformed in the external environmen t. Therefore, the manipu- lations pro duce a sense of agency (Jeannero d, 2003) for whic h the agen t p erceiv es itself as the cause of ‘in ternal actions’ (GARIM op erations) and of the effects they pro duce. Note that the concept of GARIM agency is com- patible with other concepts proposed by the litera- ture. F or example, Metzinger (2013) highlights the concept of men tal action and cognitive agency to iden tify the capacity to control own goal-directed conscious processes. Moreov er, mental action and self-con trol are concepts approached by the active inference framework (Metzinger, 2017; Hohwy and Seth, 2020). Thus, the GARIM theory captures man y asp ects of conscious goal-directed cognition 14 and agency that are considered fundamen tal b y other studies in the field. 3.5.1. A GARIM agency sc ale The GARIM agency is a suitable concept for generating a quan titative scale, which tak es in to accoun t the different levels of consciousness and flexible goal-directed b eha viours. In particular, the three features of the GARIM agency (self- mo del, emotional/p erceptual vividness, and manip- ulation control) lead to the emergence of three ‘lev- els of Consciousness’ during the expression of goal- directed b ehaviours: phenomenal consciousness, ac- cess consciousness, and manipulation consciousness (Figure 5). W e explain these three levels by de- scribing examples of human cognition. Figure 5: A scale of consciousness based on the concept of GARIM agency . Phenomenal c onsciousness piv ots on the pe- ripheral activ ations of p erceptual/emotional sub- GINPs. They are triggered b y either external p erceptual inputs or in ternal b ottom-up processes (e.g., emotional/motiv ational ev ents). The emer- gence of an unexp ected, goal-irrelev ant p ercep- tual even t is an example of this GARIM agency lev el. Indeed, while p ossibly sho wing some emo- tional/p erceptual vividness, this ev ent activ ates a temp-GINP (conscious goal-irrelev ant represen ta- tion). It is accompanied by a lo w level of GARIM agency and it is soon discarded. In case the repre- sen tation is a pre-GINP (unconscious goal-relev an t represen tation), it can b e transformed into a GINP th us leading to a higher level of GARIM agency . A c c ess c onsciousness inv olves a mild top-down selection that leads to a weak comp etition b et ween differen t sub-GINPs. This GARIM agency level can b e exemplified by the state of mind-wandering (Grub erger et al., 2011). This is a brain state, usually accompanying the p erformance of routines, that generates conscious sequential thoughts rep- resen ting temp-GINPs (e.g., thoughts on p ossible actions). In this respect, Christoff et al. (2016) sug- gest that mind-wandering inv olv es a shallow “delib- erate constrain t”, that is, a partially delib erate cog- nitiv e con trol on own thoughts. Based on our pro- p osal, this pro cess should inv olve contin uous trans- formations of pre-GINPs into GINPs and vice versa, and non-GINPs in to temp-GINPs and vice versa. These pro cesses would b e the effect of a w eak top- do wn con trol, and indeed mind-wandering can take place without aw areness (Sc ho oler et al., 2011). Manipulation c onsciousness is c haracterised by a high con trol on internal represen tations. This state is exemplified b y sp ecific forms of mindfulness ac hieved in meditation (Kabat-Zinn, 1990; Mali- no wski, 2013). F or example, fo cused meditation aims to induce a high goal-directed attentional fo- cus (e.g., on o wn breath). This amplifies the ac- cess to consciousness of goal-relev an t information (GINPs), and leads to a non-judmen tal state by strategically suppressing internal/external distrac- tions (temp-GINPs) and ruminations (T ang et al., 2015; Y ates and Immergut, 2015). Note that even in the case of unfo cused non-judgmental states (e.g., some forms of mindfulness) a higher-level goal can b e active, namely ‘to keep the whole state of med- itation in tact from distraction’. Similar features can b e shared by brain states supp orting a high at- ten tional engagement in comp etitiv e sp ort sessions (He et al., 2018; Miller and Clapp, 2011; Memmert, 2009) or intellectual games (e.g., c hess; Atherton et al., 2003; W ang et al., 2020; H¨ anggi et al., 2014). Ov erall, the GARIM agency is exp ected to con- tin uously fluctuate along the different lev els of con- sciousness. Healthy a wak e p eople migh t likely re- main most of the time within middle levels of con- sciousness, for example when carrying out daily routines (e.g., house reordering and shopping). The rest of the time they migh t ha ve transitory phases in to the lo wer levels of consciousness, and limited p eriods of time into the highest levels. The follow- ing section prop oses that there are states of con- sciousness accompanied by altered GARIM agency 15 lev els, falling b et ween the middle and the lo w lev els of consciousness. A lter e d states of the GARIM agency. The GARIM theory and the scale presented in the previous sec- tion describ e sp ecific states of consciousness during the expression of goal-directed b eha viours. Some of them may result in an altered state of GARIM agency (Figure 5). F or example, alterations of the GARIM agency could in volv e pseudo-hallucinations and hallucinations (T elles-Correia et al., 2015). Both states are experienced in the absence of ex- ternal stim uli. Ho wev er, pseudo-hallucinations are p erceiv ed as unreal dummy p erceptions whereas hallucinations are p erceiv ed as real perceptions. In- terestingly , the t wo show different levels of sensory con trollability and vividness, which are higher in pseudo-hallucinations (v an der Zwaard and P olak, 2001). These evidence are compatible with an al- teration of the GARIM agency . Dreams and lucid dreams are other conscious- ness states that could inv olve an altered GARIM agency . Dreams inv olve an uncon trolled imagina- tion during the REM sleep while lucid dreams in- v olve a partially controlled imagination (Stumbrys et al., 2012). Both states corresp ond to the genera- tion of a vivid internally simulated reality (Rev on- suo, 2006). How ev er, a higher level of control dis- tinguishes lucid dreams from dreams (V oss et al., 2009), also suggesting that a stronger activ ation of fron tal areas could cause this difference. Our pro- p osal is compatible with this evidence as the alter- ations of the GARIM agency should dep end on the influence of the top-down manipulator. 4. Comparisons of the GARIM theory with other theories The GARIM theory proposes an integrated framew ork that takes into account conscious and higher-order cognition, and thus it is compati- ble with most theories of consciousness (Seth and Ba yne, 2022). In particular, it accounts for several k ey asp ects that are considered fundamental for the emergence of conscious states (see T able 3). Here w e briefly describ e these theories and we compare them with the GARIM theory . Inte gr ate d Information the ory (IIT). The IIT (T ononi, 2008; T ononi et al., 2016; Ko ch et al., 2016; T ononi, 2004) prop oses that systems exhibit- ing high capacity of discrimination (to encode sev- eral alternative neural represen tations of cognitiv e con tents, e.g. p ercepts) and inte gr ation (to enco de sev eral differen t asso ciations b et w een differen t as- p ects of neural representations, e.g. stimuli) p o- ten tially hav e a high level of consciousness. This theory also prop oses the Φ co efficien t, a quantita- tiv e measure of the level of information in tegration. The thalamo-cortical system should hav e a key role in conscious states - th us an high Φ - due to its high synaptic in tegration and interconnection. A recent up date of the theory (Ko c h et al., 2016) has identi- fied a ‘hot zone’, lo cated within the parietal cortex, that supports the formation of conscious con tents. On the other hand, the fron toparietal system would ha ve a control role of cognitive conten ts but not a cen tral role for the emergence of a conscious state. The GARIM theory do es not delve into sp ecific asp ects of information theory , but it takes into ac- coun t key features of the I IT such as discriminabil- it y and integration. F or example, the perceptual and abstract working memory comp onen ts are ex- p ected to p erform a high ‘discrimination’ of exp e- riences. In particular, the manipulator component selects sp ecific sub-GINPs b et w een sev eral alterna- tiv e ones, th us assigning a sp ecific and stable mean- ing to experiences (high discrimination). A t the same time, the generation of stable GINPs requires a dynamic highly flexible ‘assembling’ of sub-GINPs based on suitable functional and anatomical con- nectivit y (high integration). The GARIM theory , ho w ever, has also imp or- tan t differences with resp ect to the IIT theory . First, the I IT theory lacks a functional explanation of conscious pro cesses, fundamental for dev eloping a comprehensive theory of consciousness (Cerullo, 2015). Indeed, computational systems can exhibit a high Φ while p erforming dull calculations (Seth et al., 2006; Aaronson, 2014). Second, the GARIM theory emphasises the importance of a top-down and goal-directed manipulation of represen tations, while the I IT argues that a top-down control is not fundamen tal for the emergence of conscious con- ten ts. How ever, the tw o theories may fo cus on dif- feren t conscious states. Indeed, the GARIM theory fo cuses on higher-order conscious states based on represen tation manipulation, but it exp ects the ex- istence of conscious states with a temp orary lo wer lev el of top-do wn con trol (temp-GINP). These lat- ter states app ear to b e the fo cus of the I IT theory . 16 Conver genc e-Diver genc e Zones the ory (CDZT). The CDZT (Damasio, 1989; Mey er and Dama- sio, 2009; Damasio and Meyer, 2009a) prop oses that the brain is organised on m ultiple p eriph- eral CDZs (P-CDZs; e.g., sensory cortices) and ma jor central CDZs (C-CDZs; e.g, asso ciativ e ar- eas such as prefron tal, parietal, and temp oral cor- tices). The P-CDZs transmit a b ottom-up informa- tion flo w to the C-CDZs, which p erform a top-down retro-activ ation on them. In particular, the retro- activ ation increases the meaningful integration of b ottom-up p eripheral represen tations, resulting in conscious p erception and imagination. Con versely , if the P-CDZs fail to activ ate the asso ciated pat- terns in the C-CDZs, there is no retro-activ ation and the p eripheral representations remain uncon- scious. The CDZs theory also prop oses that low- lev el somatic reactions assign an emotional v alence to the representations within the C-CDZs, giving them sufficient priorit y to enter consciousness pro- cessing (‘somatic marker hypothesis’; Bec hara and Damasio, 2005; V erdejo-Garc ´ ıa et al., 2006). A t last, the theory proposes that the represen tations at the basis of sub jective experience (C-CDZs) enco de sensorimotor relations b et ween the agent, ob jects, and even ts in the external en vironment (‘em b o di- men t approach’; Damasio and Mey er, 2009a). The GARIM theory takes into account key el- emen ts of the CDZ theory (Damasio, 1989), fur- ther specifying them with neuroscientific and com- putational details. The GARIM theory attributes a k ey role to the neural hierarc hies of the brain. Indeed, P-CDZs and C-CDZs correspond to brain structures that should supp ort p erceptual and ab- stract sub-GINPs, resp ectiv ely . F urthermore, the GARIM theory prop oses that these sub-GINPs are generated b y b ottom-up and top-down informa- tion flows. Bottom-up flows supp ort the enco ding of p erceptions in p erceptual and abstract working memories at increasing lev els of abstraction. T op- do wn flows generate sub-GINPs that are functional to the achiev emen t of goals. Both flows are con- trolled by the top-down manipulator, guided by mo- tiv ations and goals. The manipulator selects the relev an t information that trav els along cortical hi- erarc hies, thus improving the goal alignmen t of rep- resen tations. In line with the CDZT, the GARIM theory also tak es in to accoun t the role of emotions and mo- tiv ations for the assignmen t of v alence to exp e- rience. The GARIM theory sp ecifies that moti- v ational systems support sub-GINPs prioritisation through the GARIM computational operations. Fi- nally , in line with the CDZT, the GARIM the- ory emphasises the role of emotions and motiv a- tions for sub jectiv e exp erience. In particular, it prop oses that top-down manipulation pro cesses ac- tiv ate p eripheral sensory areas (imagination) and emotional/motiv ational system s (similar to the so- matic marker hypothesis; Damasio, 1989). The re- sulting activ ations then send a feedback to the cen- tral areas, asso ciating a high level of perceptual vividness and emotional v alence to sub jective ex- p erience (see Section ‘GARIM agency and the sub- jectiv e exp erience of consciousness’). Glob al Worksp ac e The ory (GWT) and Glob al Neu- r onal Worksp ac e The ory (GNWT). The GWT (Baars, 1997; Baars et al., 2003; Baars, 2005; Baars et al., 2013) proposes that consciousness relies on a set of interacting cognitive elements, which are metaphorically asso ciated with the elemen ts of a theatre: conscious conten ts (e.g., p ercepts and though ts; the ‘actors in the stage’); the glob al worksp ac e of working memory (the ‘theatre stage’); selectiv e attention (the ‘theatre sp otligh t’); exec- utiv e functions (the ‘director’); and unconscious bac kground pro cesses that interact with the global w orkspace (the ‘audience’). The theory prop oses that alternative conten ts comp ete to enter the global workspace and thereb y b ecome conscious. Selectiv e attention pro cesses, guided b y the exec- utiv e functions - in turn guided by motiv ations and goals - choose which conscious conten ts win the comp etition. The winner conten ts are br o adc aste d from the global space to the other pro cesses to sup- p ort higher-order pro cesses (e.g., decision making and self-monitoring). The GNWT (Dehaene et al., 1998a; Dehaene and Naccac he, 2001; Dehaene and Changeux, 2011) w as initially prop osed to sp ecify the neural cor- relates of the GWT based on extensiv e empiri- cal support (Mashour et al., 2020) and compu- tational formalisations (Dehaene and Changeux, 2005; Dehaene et al., 2017). The hypothesis pro- p oses the existence of tw o computational spaces in the brain. A first space is supp orted by high- densit y short/medium range connections and in- cludes many sp ecialised functional mo dules (e.g., sensory areas, motor systems, memory areas, ev al- uativ e comp onents). A second space , called ‘neu- ronal global workspace’, is supp orted b y long-range excitatory pro jections (Dehaene and Changeux, 2011) and includes a distributed set of associative 17 areas (the prefron tal, parietal, temporal and cin- gulate cortices) and cortical-sub cortical net works (e.g., the fibres of the corpus callosum and the cortico-thalamic system). This arc hitecture allows the global workspace to generate global activ ation patterns with v ariable duration ( ignitions ), inv olv- ing distributed interconnected net works. These patterns strongly comp ete and inhibit or fa vour re- lated patterns within p eripheral specialised mod- ules (e.g., p erceptions, emotions and actions related to an ob ject). The fron tal-parietal system pla ys a k ey role in supp orting this top-down amplification of information. The biological underpinnings of the GNWT hav e b een extended to envisage the exis- tence of ‘buffers’ (working memories) betw een the sensorial cortices and the neuronal workspace (Raf- fone et al., 2014b, 2015). At last, the GNWT has re- cen tly b een integrated with inferential frameworks (Mashour et al., 2020), suggesting that top-down amplification corresp onds to an inferential pro cess applied on b ottom-up sensory inputs. The GARIM theory in tegrates the main con- cepts of the GWT (Baars, 1997) and the GNWT (Dehaene and Changeux, 2011). In addition, it enric hes those concepts b y sp ecifying the p ossi- ble goal-directed computations (e.g., manipulation functions) and the brain mec hanisms that might underlie them. First, as the GWT and GNWT, the GARIM theory assumes a ‘centre-periphery’ arc hitecture underlying conscious states as well as goal-directed b eha viours. In particular, it prop oses m ultiple p erceptual working memories that trans- mit information to the abstract working memory . Therefore, it integrates such information and dis- patc hes the result back to the p eripheral struc- tures (the ‘broadcasting’ of GWT and GNWT). Second, the mec hanisms underlying the generation of GINPs are compatible with those supp orting ‘ig- nitions’. Indeed, an ignition is a coheren t activ ation of link ed lo cal neural patterns in central and p eriph- eral areas. Third, the GARIM theory ascrib es a key role to the fron to-parietal brain system, prop osing that it is fundamental for the top-down and goal- directed control of sensorimotor cortical pathw a ys. While sharing these imp ortan t elemen ts with the GWT and GNWT, the GARIM theory fur- ther sp ecifies them. First, in the GWT the pat- terns activ ated by ignitions are mainly generated b y p ercepts. Instead, the GARIM theory p ostu- lates that the volitional goal-directed generation of GINPs dep ends on the selection of sub-GINPs b y the top-do wn manipulator. Second, while assign- ing an imp ortant role to the cortical fronto-parietal system, the GARIM theory highlights the pivotal role that the basal ganglia-thalamo-cortical system pla ys in the manipulation of sub-GINPs. Finally , the GARIM theory sp ecifies the functioning of the b ottom-up and top-do wn information flows in terms of computational manipulation op erations (abstrac- tion and sp ecification/generativ e mechanisms). Higher-or der the ories (HOTs). HOTs represent a family of theories originally formulated in philos- oph y (for a review, see Brown et al., 2019). All HOTs share the idea that first-or der r epr esenta- tions , for example the activ ation of patterns within the early stages of the visual cortex, are necessary but not sufficient to ha ve a conscious exp erience. In particular, an agen t can generate conscious con- ten ts only after first-order states hav e b een ev alu- ated and meta-represented by higher-or der r epr e- sentations . The Radical Plasticit y theory (Cleere- mans, 2007, 2011), an instance of the HOTs, pro- p oses that meta-represen tations show three sp ecific features: robustness, stability and distinctiveness. The theory has been recently integrated with in- feren tial pro cesses (Cleeremans et al., 2020). Most HOTs suggest that a certain level of ‘inner aw are- ness’ of one’s ongoing mental pro cesses is necessary to ha v e consciousness. The claims of the HOTs ha ve b een supp orted by empirical evidence, high- ligh ting the contribution of frontal netw orks in the formation of conscious higher-order representations (Lau and Rosenthal, 2011). At last, the HOTs prop ose that first-order and second-order represen- tations inv olve the interaction b et ween sub cortical and cortical systems, leading to an explanation of emotional asp ects of conscious exp erience (LeDoux and Brown, 2017). The GARIM theory sp ecifies the k ey concepts of the HOTs (Brown et al., 2019) in terms of computa- tional brain mec hanisms. First, the GARIM theory prop oses that the in teraction of four comp onents leads to the encoding and selection of sub-GINPs at increasing lev els of abstraction. The abstract sub- GINPs hence integrate the conten ts of low er-level p erceptual sub-GINPs at a more abstract level, thus represen ting a form of meta-representations. More- o ver, in agreement with the Radical Plasticity The- ory (Cleeremans, 2011), GINPs should exhibit the three key features of robustness, stability and dis- tinctiv eness b ecause they tend to (a) enco de dis- tinctiv e elements of goal-directed pro cesses and (b) remain stable ov er time as long as they are relev an t 18 for the set goal. On the other hand, unconscious represen tations (e.g., non-GINPs) can briefly access consciousness (temp-GINPs) but then quickly fade a wa y (lo w stability). Finally , the GARIM theory can also account for the ‘inner a wareness’ p ostu- lated by HOTs. In particular, the goal-directed in- ternal manipulation of representations give rise to a sense of agency that can accompany inner aw are- ness (see Section ‘Sub jective experiences during conscious goal-directed b eha viours: the GARIM agency’). Sensori-motor the ory (SMT). The SMT prop oses that conscious exp erience piv ots on the interactions b et w een the brain, the b ody , and the environmen t (O’Regan and No e, 2001; O’Regan et al., 2005). The theory was developed within the theoretical framew orks of emb o die d c o gnition (Anderson, 2003; Garbarini and Adenzato, 2004; Borghi and Cimatti, 2010) and enactivism (Hutto, 2005). The theory substan tially div erges from the other theories as it de-emphasises the role of brain pro cesses and rep- resen tations, highlighting instead the imp ortance of sensorimotor exp erience. The theory prop oses that sensorimotor c ontingencies (the even ts linking ac- tions to sensory changes; Jacquey et al., 2019) are fundamen tal in determining the phenomenal sen- sations that accompan y conscious exp erience. Dif- ferences in these sensorimotor activities distinguish sensory exp erience and reasoning/imagination pro- cesses. In particular, sensory exp erience has ‘alert- ness’ - the capacity to exogenously attract our at- ten tion - and ‘corporality’ - the fact that b odily actions immediately mo dify the sensory input. In agreement with the SMT (O’Regan and No e, 2001), the GARIM theory supp orts the idea that consciousness pla ys a fundamen tal function for adaptation. How ever, the SMT prop oses that the k ey function of consciousness is the generation of a close coupling b et ween motor action and its per- ceiv ed effects. Instead, the GARIM theory prop oses that the k ey function of consciousness is to enhance goal-directed processes to increase b eha vioural flex- ibilit y . Moreo ver, the SMT pushes the embo died view of cognition tow ards anti-represen tationalist p ositions (Pennartz, 2018). The GARIM theory departs from these p ositions as ‘represen tations’ and ‘manipulation of representations’ are key con- cepts for it. Ho wev er, k ey theoretical asp ects at the basis of b oth theories hav e b een recently rec- onciled b y highlighting that goal represen tations could supp ort a link b et ween actions and their p er- ceiv ed effects (sensory-motor contingencies) (Bal- dassarre et al., 2018; Jacquey et al., 2019; Mannella et al., 2018). Finally , in line within the SMT, the GARIM theory clearly emphasises the imp ortance of agency for the generation of sub jective conscious exp erience. Ho wev er, the GARIM agency is cen- tred on goal-directed representation manipulations while the SMT is fo cused on sensory-motor inter- action with the environmen t. Pr e dictive Pr o c essing the ories (PPTs). The PPTs (for a review, see Hohwy and Seth, 2020) are a fam- ily of theories that link conscious states to the con- cepts of ‘predictiv e co ding’, ‘error minimisation’, and ‘world mo del’. According to the theory , the brain implemen ts internal world mo dels based on stac ked dynamic neural lo ops. At eac h lo op, the higher lev els pro duce predictions ab out the acti- v ation of the low er levels (hence ‘predictiv e cod- ing’), which in the low est lo ops directly predict the p ercepts of the world. On the other hand, the low er lev els flow information up ward and com- pute prediction-errors b y comparing the top-down predictions and their b ottom-up activ ations. The prediction errors supp ort a p erp etual refinement of the w orld mo dels. The Active Inference F rame- w ork (AIF), an instance of PPTs, proposes that the prediction-error can b e minimised also by p erform- ing actions (e.g., o cular mo vemen ts) to pro duce exp ected sensory data (predictiv e con trol). Over- all, the brain correlates of PPTs corresp ond to the b ottom-up hierarchical flows, for example from sen- sory cortices to prefron tal systems, and top-do wn feedbac k flo ws, from higher-order brain systems to lo wer sensory areas. The GARIM theory shares some important el- emen ts with the PPTs. Both theories highlight the imp ortant role for consciousness of bidirectional brain hierarc hies. In particular, the GARIM theory prop oses that top-do wn information flo ws along the hierarc hies implement generative pro cesses recon- structing representations at low er-levels. This pro- cess is fully in line with the generative mec hanisms of predictive co ding. In addition, b oth the GARIM and the AIF explicitly refers to an active top-down con trol of action. Although these common p oin ts, the GARIM the- ory has additional elements and some div ergen t p o- sitions. First, the GARIM top-down manipulator p erforms several goal-oriented op erations on kno wl- edge, at differen t abstraction lev els. These pro- cesses generate new kno wledge not only by inter- 19 p olating previously acquired knowledge, but also b y extrapolating it to pro duce more creative de- vian t represen tations (e.g., imagining a new to ol). Instead, the PPTs and AIF, piv oting on the mech- anism of prediction error, are more closely linked to in terp olation pro cesses. Second, the GARIM the- ory ascrib es a cen tral role to goal-directed cognition and b eha viour while initial PPTs prop osals did not do so. How ev er, recent prop osals of the AIF hav e started to interpret goal-directed pro cesses (Pez- zulo et al., 2015; Matsumoto et al., 2022; Hohwy and Seth, 2020; F riston et al., 2016), thus pro duc- ing a p oten tial common ground with the GARIM theory . A t last, although the GARIM theory as- crib es a key role to information flows in cortex, it also prop oses that basal-ganglia and cortical selec- tion mec hanisms play a k ey role to instantiate the manipulator’s op erations. Neur or epr esentationalism the ory (NR T). The NR T (P ennartz, 2015, 2018, 2022) is a theoretical frame- w ork that defines Consciousness as a ‘multimodal and situational survey’. It proposes that con- scious states dep end on multimodal/multi-lev el rep- resen tations, which are fundamental to sub-serv e goal-directed b eha viours (e.g., planning). This framew ork prop oses five features that describ e con- scious exp erience: ‘multimodal richness’, the emer- gence of sensations in multiple distinct modali- ties; ‘situatedness/immersion’, the sensation that our b ody is immersed in the space and has a cen- tral p osition with resp ect to the surrounding stim- uli; ‘unity/in tegration’, the emergence of a sin- gle undivided and multi-modal representation; ‘dy- namics/stabilit y’, the emergence of dynamic p er- ceptions (e.g., external environmen t c hanges) and static p erceptions (e.g., stationary ob jects); ‘in ten- tionalit y’, the generation of signals that are in- terpreted as something other than ourselv es. At last, the NR T highlights that predictive pro cessing is a suitable framework for describing the neuro- computational basis of conscious states. In par- ticular, it prop oses that multi-lev el representa- tions emergently lead to multimodal and spatially wide ‘sup er-inferences’, corresp onding to phenome- nal exp eriences. The GARIM theory and the NR T share the idea that conscious states hav e the scop e to supp ort goal-directed b eha viours. Notably , they are the only t wo theories that explicitly and systemati- cally prop ose this bridge. Moreo ver, the t wo the- ories share some hallmarks that describ e conscious states. In particular, some features of conscious represen tations prop osed by the NR T are consis- ten t with the definition of GINPs. F or example, GINPs are defined as in tegrated systemic represen- tations whose sub-parts encode different asp ects of goal-directed pro cesses (e.g., motiv ations, p ercep- tions, actions). Moreov er, GINPs are formed by sub-GINPs at differen t lev els of abstraction, from mo dalit y-sp ecific working memories to multi-modal abstract w orking memories. At last, GINPs are ‘em b o died representations’ that integrate asp ects of the en vironment, the agent and their relation- ships (e.g., action outcomes). Therefore, the ‘multi- mo dal/m ultilev el representations’ prop osed by the NR T partially ov erlaps with the concept of GINPs. Despite these commonalities, the GARIM theory sho ws key differences with the NR T. First, GINPs and sub-GINPs are characterised b y a sp ecific di- mension defined ‘goal-relatedness’ (i.e. their relat- edness with resp ect to the set goal), not considered b y the NR T. Second, the tw o theories tend to fo cus on tw o different asp ects of goal-directed b ehaviour. In particular, the NR T fo cuses on the emergence of the b est representations which then subserve goal- directed pro cesses. Instead, the GARIM theory mostly fo cuses on the representation manipulation op erations that constitute goal-directed pro cesses and b ehaviour. 4.1. Inte gr ating key asp e cts of Consciousness into a neur o-functional fr amework of flexible go al- dir e cte d b ehaviour. Recen t w orks prop ose an analysis and compar- ison of the main theories of consciousness (Seth and Bayne, 2022; Del Pin et al., 2021). Ab o ve we ha ve compared these theories with the GARIM the- ory , highlighting their similarities and differences (for a summary , see T able 3). Due to its func- tionalist systemic approac h, the GARIM theory con tributes to integrate the other theories on con- sciousness at tw o levels: a ‘bac kground integration’ lev el and a ‘focused integration’ lev el. Concerning the first level, multiple concepts from previous the- ories describ e ‘brain functioning/organisation prin- ciples’ with whic h the GARIM theory is compatible (e.g., ‘information integration/discrimination’, ‘em- b odiment and sensory-motor contingencies’, and ‘predictiv e inferen tial pro cesses’). Concerning the second level, some concepts proposed b y other the- ories are cen tral also for the GARIM theory (e.g., ‘first/second order represen tations and inner a ware- ness’, ‘hierarchical bidirectional flows’, ‘broadcast- 20 Theories Key concepts Information integration and discrimination Hierarchical bidirectional flows Broadcasting, Ignitions First/second order representations, Inner awareness Embodiment, Sensorimotor contingencies Predictive inferential processes Multi-modal and multi-lev el situated representations Goal-aligning representation manipulations IIT ✓ ✓ ✗ ✗ ✗ ✗ ✗ ✗ ✗ CDZT ✗ / ✓ ✓ ✓ ✗ ✗ ✓ ✗ ✓ ✗ GWT/GNWT ✗ ✓ ✓ ✓ ✗ ✗ ✓ ✗ / ✓ ✗ / ✓ HOTs ✗ ✗ / ✓ ✗ ✓ ✓ ✗ ✗ / ✓ ✗ ✗ SMT ✗ ✗ ✗ ✗ ✓ ✓ ✗ ✗ ✗ PPTs ✗ / ✓ ✓ ✗ ✗ ✗ / ✓ ✓ ✓ ✗ / ✓ ✗ NR T ✗ / ✓ ✓ ✗ ✗ ✗ / ✓ ✓ ✓ ✓ ✗ GARIM theory ✓ ✓ ✓ ✓ ✓ ✓ ✓ ✓ ✓ T able 3: Main concepts of the theories of consciousness considered in this work. Symbols: ✓ ✓ : concept pivotal for this theory; ✓ : concept compatible/encompassed b y this theory; ✗ / ✓ : concept partially compatible/encompassed b y this theory; ✗ : concept not compatible/encompassed by this theory . ing/ignitions’, and ‘m ulti-mo dal/m ulti-level situ- ated representations’). Ov erall, the GARIM theory highlights that higher-order cognition and consciousness necessar- ily require all these elemen ts. Ho wev er, the tw o dif- feren t levels of integration could differentially b en- efit scientific and technological fields. F or example, elemen ts of the fo cused integration could aid the design of computational mo dels. Indeed, they usu- ally repro duce sp ecific functions and neural mec ha- nisms to explain brain and b eha viour. On the other hand, elements drawn from background in tegration could aid the design of AI/rob otic architectures. In- deed, these systems can also b enefit from algorith- mic solutions that are only conceptually inspired b y higher-order human cognition and consciousness (see section ‘Implications of the GARIM theory for computational mo delling, AI and Rob otics’). 5. Experimental and clinical implications of the GARIM The GARIM theory represents a theoretical framew ork that has implications for several fields. In this section we first consider its contribution to the understanding of the concept of ‘Intelligence’. Then we prop oses interpretations of psyc hological and neuropsyc hological evidence on goal-directed b eha viours and consciousness. 5.1. GARIM the ory and Intel ligenc e The GARIM theory fo cuses on the higher-order goal-directed cognition inv olving conscious states. These same pro cesses could be at the basis of the expression of ‘intelligen t pro cesses and b eha viours’. Although the inv estigation of intelligence is b ey ond the scop e of this w ork, the GARIM theory can con- tribute to its understanding, in particular to clarify its relationship with flexible goal-directed cognition and consciousness. The term ‘intelligence’ refers to a comp osite con- struct encompassing multiple areas of competence (Gardner, 2000) and is measured with different scales of in telligence (e.g., W AIS; Benson et al., 2010). Recently , new theoretical framew orks hav e stressed the difference b et w een domain-general and domain-sp ecific intelligence (Burk art et al., 2017), also strengthening the relationship b et ween in tel- ligence and goal-directed b eha viour (Chiapp e and MacDonald, 2005; T egmark, 2017). In our previous computational proposals we mo d- elled the interaction b et ween domain-general pro- cesses (e.g., working memory and motiv ational sys- tems) and domain-specific competence (e.g., sen- sory and motor learning). This allow ed the study of task-related representation learning (Granato et al., 2022b) and goal-directed representation manipu- lation (Granato and Baldassarre, 2021; Granato et al., 2020, 2022a; Granato et al.). On the basis of these works, we explicitly prop osed the idea that the flexibility characterising domain-general in tel- ligence rests on the goal-directed manipulation of represen tations (Baldassarre and Granato, 2020). The GARIM theory extends these ideas to higher- order cognition and consciousness. In particular, it prop oses that consciousness b oosts flexibility , a key 21 asp ect of general-domain intelligence. This flexi- bilit y might aid the acquisition of domain-specific comp etences (e.g., motor skills) through the top- do wn guidance of the learning pro cesses. F urther- more, flexibilit y migh t supp ort the on-shot selec- tiv e p erformance of previously acquired automatic b eha viours. This prop osal is compatible with some imp or- tan t features of other theories of consciousness. F or example, the Radical Plasticit y Theory (belonging to the HOTs) suggests that consciousness b oosts learning pro cesses. Moreo ver, the GWT and the GNWT suggest that the global-workspace informa- tion broadcasting improv es the lo cal learning of rep- resen tations within p eripheral brain sub-mo dules (e.g., motor mo dules). Moreov er, the prop osal is compatible with the concept of information in tegra- tion prop osed by the IIT. Indeed, flexible in telligen t b eha viour should require a high information inte- gration within higher-order brain areas (e.g., the abstract working-memory), in turn influencing the lo wer-order ones (e.g., the motor and perceptual ar- eas). 5.2. An interpr etation of exp erimental and clinic al evidenc e b ase d on the GARIM the ory The GARIM theory may b e useful in in terpreting psyc hological and neuropsyc hological evidence on goal-directed b eha viours and consciousness. F ur- thermore, it ma y stimulate the developmen t of new exp erimen tal paradigms inv estigating the func- tional role of conscious states in flexible cognition. 5.2.1. L esion studies, go al-dir e cte d c o gnition, and c onscious states The relationship b et ween brain lesions and con- sciousness disorders is still not fully clear. In par- ticular, there is no research that systematically links impairments of fron tal systems and basal ganglia, whic h play a key role for our prop osal, with consciousness disorders. How ev er, the em- pirical supp ort of HOTs indicates that PF C le- sions cause a deficit in consciousness-related pro- cesses (e.g., metacognitiv e capabilities; Lau and Rosen thal, 2011). Moreo ver, recen t prop osals sug- gest that PFC lesions could influence consciousness in unnoticeable wa ys (F ox et al., 2020). On the other hand, v arious studies show that basal gan- glia lesions cause a general consciousness impair- men t (e.g., Rohaut et al., 2019). Moreov er, a bulk of studies (Ell et al., 2006, 2010; W ard et al., 2013; Price et al., 2009) show that fo cal damages of basal ganglia impair explicit/conscious reasoning but not implicit/unconscious categorisation. These studies do not explicitly inv estigate consciousness, but they put in relation impairments of k ey elemen ts of goal- directed cognition and explicit/conscious processes. In general, the GARIM theory do es not pro- p ose a conclusiv e explanation regarding the relation b et w een consciousness disorders and frontal/basal ganglia lesions. How ev er, it prop oses a link be- t ween these lesions, the p ossible alterations of explicit/conscious cognitive pro cesses and goal- directed flexible b ehaviours. F or example, the GARIM theory predicts that extended lesions to PF C systems and asso ciativ e p ortions of basal gan- glia would impair abstract working memory and the top-do wn manipulator. Their impairment should corrupt the manipulation of GINPs. In particu- lar, this alteration should lead to a reduced abil- it y to transform pre-GINPs in GINPs or to sup- press temp-GINPs (e.g., distractors). Therefore, these alterations should mak e an agent less gen- erativ e and fo cused. These predictions are consis- ten t with the clinical literature on goal-directed b e- ha viour. Indeed, alterations of PFC and basal gan- glia cause cognitive inertia, namely a reduced ca- pacit y to inten tionally generate/activ ate strategies required to successfully complete a given program of actions (Levy, 2012, 2021). Despite these studies do not explicitly refer to consciousness, our theory prop oses a link b et w een conscious states and the ab o v e deficits. Indeed, cognitive inertia should alter the generation of GINPs, th us the exploitation of conscious and generative pro cesses to express flexi- ble b eha viours. Moreov er, the impairment of these structures could alter the GARIM agency , explain- ing the emergence of hallucinatory perceptual rep- resen tations after fron tal and basal ganglia lesions (F ornazzari et al., 1992; F rith, 1996; W o darz et al., 1995; McMurtray et al., 2014). Note that these alterations do not corre- sp ond to global alterations of consciousness (vigi- lance/a wareness). Indeed, the GARIM theory pre- dicts that a fo cused lesion of PFC systems and basal ganglia would not cause a general loss of conscious- ness (e.g., coma). Moreov er, they w ould not pre- v ent the access of stimuli to consciousness (phenom- enal consciousness). In summary , the GARIM theory predicts that the fron tal cortex and basal-ganglia impairments alter the link b etw een consciousness and flexi- ble b ehaviours. In particular, they imp ede an 22 adequate emergence/managemen t/manipulation of GINPs (access consciousness and manipulativ e con- sciousness). This corresp onds to an inefficient ex- ploitation of conscious processes for generating new kno wledge and new p erspectives during the expres- sion of goal-directed b eha viours (e.g. problem solv- ing). 5.2.2. Exp erimental evidenc e: the pr e dictions of the GARIM the ory The GARIM theory do es not yet ha ve direct em- pirical supp ort, but it pro duces sp ecific exp erimen- tal predictions. Imp ortantly , these predictions are in line with the exp erimental evidence provided by other theories of consciousness. First, the GARIM theory predicts that p ercep- tual sub-GINPs, inv olving the posterior higher- order sensory cortices, should remain activ e throughout the p erformance of explicit tasks. These activ ations should supp ort b ottom-up ab- straction and top-down generative pro cesses. This prediction matches the exp erimen tal evidence at the basis of the I IT. Indeed, by con trasting stim- ulation effects during coma and wak efulness, ev- idence sho ws that a sustained activ ation of the p osterior ‘hot-zone’ is necessary for consciousness (Ko c h et al., 2016). Second, the GARIM theory also predicts that the emergence of GINPs is preceded by the activity of the top-do wn manipulator, inv olving the syner- gistic activ ation of the fron to-parietal system and the basal ganglia. This prediction agrees with the evidence pro duced by the GNWT on contrastiv e tasks (e.g., masking, bino cular riv alry , attentional blinking; Aru et al., 2012), highligh ting that con- sciousness emerges due to a strong activ ation of the fronto-parietal areas (‘ignitions’; Dehaene et al., 2011). The activ ation of the top-down manipulator and the emergence of GINPs w ould correspond to the ignition pro cesses recorded in these studies. In addition, the GARIM theory further predicts that, giv en the same stimuli, different ignitions (GINPs) w ould emerge when differen t goals are pursued. Third, some studies argue that there can b e a dis- so ciation b et ween attention and explicit/conscious pro cessing (Ko ch and T suc hiy a, 2007). These pro- p osals are usually link ed to bottom-up attention rather than top-do wn attention. Indeed, atten- tion proce sses are generally considered ‘necessary’ to pass from unconscious to conscious pro cessing (V an Bo xtel et al., 2010; Raffone et al., 2014a; Pitts et al., 2018), but they may not be ‘sufficien t’. In this resp ect, the GARIM theory predicts that: (a) stimuli having a high relev ance for the pursued goals hav e a higher c hance to b e selected by atten- tion and thus to access consciousness (pre-GINPs); (b) stimuli with a high b ottom-up saliency may b e able to en ter consciousness (temp-GINPs) but they fade in case of a lack of supp ort from top-down goal-directed mechanisms. Finally , the GARIM theory predicts that a basal- ganglia/prefron tal cortex activ ation is necessary to generate a goal representation. This prefrontal ac- tiv ation precedes and guides the GINP generation and conscious goal-directed behaviour. This predic- tion agrees with evidence reported b y the HOTs. In particular, these show that a prefron tal activ ation is necessary to supp ort second-order activ ations and the ev aluation of own knowledge (Lau and Rosen- thal, 2011). Our prop osal agrees with these in ter- pretations, as GINPs inv olve second-order represen- tations integrating p erceptual, motiv ational, and motor representations. Moreov er, the GARIM the- ory sp ecifies that conscious processes in volv e b oth the manipulation of representations and the ev alu- ation of their alignment with the pursued goal. Ov erall, how ev er, we b elieve that the tasks on consciousness prop osed so far can only partially test the basic principles of the GARIM theory . The next section elab orates on this idea. 5.2.3. T owar ds new tasks and pr oto c ols that test GARIM the ory mor e dir e ctly Not withstanding the growing evidence, empirical supp ort of the ma jor theories of consciousness is still unsatisfying (Y aron et al., 2022; Del Pin et al., 2021; Do erig et al., 2021; Melloni et al., 2021). The GARIM theory can con tribute to identify the prob- lems that prev ent the collection of more solid em- pirical evidence on consciousness. Common exp erimen tal proto cols (e.g., con- trastiv e metho ds; Aru et al., 2012) mostly fo cus on the first stage of conscious processing consid- ered by the GARIM theory , requiring exp erimen tal participan ts to dete ct a stimulus and to p erform simple actions in resp onse to it (e.g., reply ‘yes/no’ or c ho ose one b et ween few options, e.g. by voice or b y pressing buttons). According to the GARIM theory , these tasks are not sufficient to test manip- ulativ e consciousness. In particular they fo cus only on awar eness , explained b y the GARIM theory as the initial passage from non-GINPs/pre-GINPs to temp-GINPs (phenomenal consciousness). Instead, 23 these tests are not sufficient to dissociate phenom- enal consciousness and manipulativ e consciousness, the latter of which requires sustained manipulative pro cesses. Indeed, exp eriments capable of mak- ing this distinction should in v olve new goals or new conditions that require goal-directed pro cesses (planning or problem-solving). Alternativ ely , they should require the re-ev aluation of relationships di- rected to increase goal-alignment, for example in relation to action-subgoal or subgoal-goal relation- ships. F or example, W eiskran tz (1995) discussed a p ossible exp erimental approach potentially testing goal-directed conscious processes. The author con- siders how blindsigh t patien ts can successfully dis- criminate stimuli without aw areness (P¨ opp el et al., 1973; W eiskrantz, 2004). Moreov er, paraplegic pa- tien ts can pro duce lim b resp onses again without a wareness (W eiskran tz, 1991). With b oth these pa- tien ts, ‘commentary actions’ (e.g., ‘press a button’ or ‘verbally rep ort y our exp erience’) are necessary to chec k the presence of aw areness. Similarly , to test these pro cesses in animals it is necessary to pre- train them in the use of commentary actions (e.g., press a button; Co wey and Sto erig, 1995). The key p oin t is that b oth h umans’ and animals’ commen- tary actions migh t in v olve habitual processes rather than in tentional conscious pro cesses. T o av oid this problem, new exp erimen tal paradigms ha ve b een dev elop ed to explicitly test the presence of goal- directed pro cesses (e.g., dev aluation; Balleine and Dic kinson, 1998; Mannella et al., 2016). This pro- p osal supports the idea that an effectiv e exp eri- men tal v erification of goal-directed manipulations of representations can b e a key step to c heck the op eration of consciousness. W e started to inv estigate the concept of repre- sen tation manipulation with computational mo d- els (Granato et al., 2020; Granato and Baldassarre, 2021; Granato et al., 2022a; Granato et al.) b y us- ing the Wisconsin Card Sorting test (Heaton et al., 1993). Ev en if this test measures executive func- tions and not consciousness, it inv olv es an explicit categorisation and requires imp ortan t representa- tion manipulation pro cesses (e.g., the selection of differen t representations to b est support a flexi- ble goal-direct b eha viour in a changing environ- men t). Despite its relev an t features, how ever, the test is not yet able to chec k v arious asp ects of con- sciousness considered relev ant by the GARIM the- ory (e.g., multi-stage planning or problem solving). Ov erall, adequate tasks testing the GARIM pro- cesses should complement existing paradigms fo- cused on testing aw areness. In particular, an ideal task should hav e these elemen ts: (a) test p er- ceptual awar eness , for example require to iden- tify/categorise input patterns based on explicit rules; (b) request the achiev ement of new go als , face new c onditions , or impr ove go al-alignment so as to require the in ternal manipulation of repre- sen tations to pr o duc e new know le dge ; (c) test the sp ecific use of the GARIM manipulation op er ations (abstraction, sp ecification, decomp osition, comp o- sition); (d) test the pro cesses of monitoring of go al- alignment ; (e) test the key elemen ts of the GARIM agency: self-mo del, emotional/p er c eptual vividness, and manipulation c ontr ol . 6. Implications of the GARIM theory for computational mo delling, and for AI and Rob otics The GARIM theory tak es into account both neu- ral and computational asp ects of conscious and goal-directed behaviour. Indeed, it has b oth sci- en tific and tec hnological implications. First, the theory pav es the wa y to the dev elopment of new computational mo dels. In particular, these could capture computational op erations at the basis of conscious and flexible goal-directed b eha viour (e.g., top-do wn manipulation) and related neural mecha- nisms (e.g., competitive cortical and sub-cortical se- lection mec hanisms). These mo dels could pro duce quan titative predictions to b e tested against sp e- cific empirical data, thus corrob orating our theory . Second, the theory provides a guideline to p ossibly enhance the current AI and rob otic systems. These systems might b e emp o wered with functions and comp onen ts proposed by the GARIM theory . The upgraded systems should b e ev aluated for their abil- it y to impro ve p erformance with resp ect to current systems (e.g., in terms of goal-oriented flexibility and learning sp eed). 6.1. T owar ds c omputational mo dels of the GA RIM the ory W e already op erationalised the three-comp onen t theory with a computational mo del (Figure 6; Granato and Baldassarre, 2021; Granato et al., 2020, 2022a; Granato et al.). This mo del is sup- p orted b y a neuro-inspired system arc hitecture based on machine learning elements (generative mo dels, recurren t neural netw orks, and reinforce- men t learning) and nov el brain-inspired algorithms. 24 The mo del was v alidated with human exp erimen- tal data in v arious conditions (e.g., frontal pa- tien ts, P arkinson, Autism) and ages (e.g., chil- dren, teenagers, young adults, and middle adults). In particular, the mo del repro duced data from man y cohorts of human participants p erforming the Wisconsin card sorting test (W CST; Berg, 1948; Heaton et al., 1993). W e used the WCST to test the model b ecause, although it was initially pro- p osed to test executive functions in general, it has no w b ecome the most commonly used neuropsy- c hological test of cognitive flexibility (Miles et al., 2021). In this respect, the test requires a top-down switc hing of in ternal representations to successfully accomplish a goal when the environmen t changes (Granato and Baldassarre, 2021). Figure 6: Schema of an already published computational model of the three-component theory (Granato and Baldas- sarre, 2021; Granato et al., 2020, 2022a; Granato et al.). The model is a starting p oint for buildi ng GARIM-inspired com- putational models. Ov erall, the mo del emulates h uman flexible goal- directed cognition and b eha viour. Since it is based on the three-component theory , it emulates three of the four comp onen ts p ostulated by the GARIM the- ory (a hierarchical p erceptual system, an executiv e w orking memory , and a top-do wn manipulator), sensory-motor lo ops, and first-order/second order represen tations and manipulations. Although the mo del sp ecifically aimed to solve the WCST and did not consider conscious processing, it could still cap- ture the C1 and C2 lev els of simulation prop osed b y mac hine consciousness (Gamez, 2008). In particu- lar, the mo del shows an explicit rule-based categori- sation process relev ant for consciousness functions. Moreo ver, the mo del presents v arious architectural and functional elements supporting consciousness in the brain. F or these reasons, this mo del rep- resen ts a p ossible starting p oin t for building new computational mo dels following the principles of the GARIM theory . T o this end, w e now prop ose a ‘blueprin t architecture’ giving guidance to this pur- p ose (Figure 7). The architecture should b e supp orted by an ade- quate in teraction of the four k ey comp onen ts (p er- ceptual hierarchies, working memory , manipulator, and motiv ation comp onen ts). How ev er, their im- plemen tation could follow different approac hes that em ulate the brain mec hanisms at different levels of detail. F or example, the mo dels could b e imple- men ted with neuro-inspired algorithms (e.g., neural net works and reinforcemen t learning metho ds) ab- stracting from the details of the brain mechanisms (as w e done in Granato and Baldassarre, 2021). On the other hand, the mo dels could emulate finer bi- ological details of the brain (e.g., spiking neurons and neuronal connectivit y; Dehaene and Changeux, 2005; Day an and Abb ott, 2001). The following paragraphs examine p otential approac hes to imple- men t each comp onen t of blueprin t architecture. 6.1.1. Per c eptual c omp onent: gener ating p er c eptual and higher-or der GINPs The p erceptual hierarchical comp onent should be able to p erform b oth abstraction and sp ecification based on generative mechanisms. Deep Belief Net- w orks (DBNs; Hinton et al., 2006; Hinton, 2012) are suitable for implementing this function. They can learn input representations at increasing lev- els of abstraction based on statistical regularities and task demands (Granato et al., 2022b). More- o ver, they are able to generate representations on the basis of previous inputs and top-do wn gener- ativ e pro cesses (Granato and Baldassarre, 2021). Spiking-neur on neur al networks are another ap- proac h that can b e used to p erform represen tation learning of k ey elements and timed chains (Kap- p el et al., 2014). These metho ds can also b e used to implemen t world mo dels enco ding sequences of w orld states within planning arc hitectures (Rueck- ert et al., 2016; Basanisi et al., 2020). Pr e dictive c o ding is another suitable approach to implemen t this function (Rao and Ballard, 1999; Pezzulo, 2014; Donnarumma et al., 2017). In this resp ect, recent approac hes integrate predictive co ding with goal- orien ted systems (P ezzulo et al., 2015; Matsumoto et al., 2022; Jung et al., 2019a). A key asp ect of computation mo dels of the GARIM theory in volv e the mechanisms used to 25 Figure 7: The GARIM blueprin t architecture aiding the realisation of specific implemen tations of mo dels follo wing the GARIM theory principles. Italics in brack ets: main brain neural mechanisms (structure and processes) p ossibly implementing the components. supp ort the encoding and dynamics of sub-GINPs and GINPs. The activ ation of sub-GINPs could rely on local neural biased comp etitions taking place at differen t levels of abstraction (e.g., as mo delled in competitive neural circuits and self- organising maps; Mysore and Kothari, 2020; Koho- nen, 2001; Diehl and Co ok, 2015). The generation of GINPs could rely on lo cal winning p opulations, enco ding sub-GINPs, that could excite other win- ning p opulations in distal areas through long-range excitatory connections (e.g., as mo delled in Miikku- lainen et al., 2006). Neural mechanisms analogous to these ha v e b een already used in models prop osed within the GNWT Dehaene and Changeux, 2005. 6.1.2. Working memory c omp onent: the long- lasting activation of GINPs The working memory comp onen t should supp ort the long-lasting activ ation of GINPs in the absence of their initial in ternal and external triggers. R e cur- r ent Neur al Networks (RNNs; Barak and Tso dyks, 2014) are suitable mo dels to em ulate these func- tions. Indeed, they emulate the dynamic re-en trant circuits of PF C systems and basal ganglia-thalamo- cortical lo ops. A t the same time, basal ganglia- lik e selection mechanisms of the manipulator could upload/do wn information from suc h recurren t cir- cuits (e.g., OReilly and F rank, 2006; Holcman and Tso dyks, 2006). R eservoir c omputing (for a review see Luk o ˇ sevi ˇ cius and Jaeger, 2009) is another suit- able approac h to implement dynamic w orking mem- ories. In particular, it exploits recurren t sto c hastic net works of whic h activit y is ‘read-out’ by exter- nal units. These units pro ject bac k to the recur- ren t netw orks, learning to induce in them the de- sired dynamic pattern. Reservoir netw orks are suit- able to emulate different details of the brain, indeed they can b e implemented with firing-rate neurons (e.g., ‘ec ho-state netw orks’; Jaeger, 2001) or spiking neurons (e.g., ‘liquid state machines’; Maass et al., 2002). 6.1.3. Manipulator c omp onent: sele ction me cha- nisms sculpting GINPs The manipulator comp onen t should b e able to implemen t the GARIM op erations, thus sculpting 26 GINPs to generate knowledge. Computational ap- proac hes that emulate the functioning of the basal ganglia-thalamo-cortical lo ops could b e a starting p oin t (Schroll and Hamk er, 2013). In particular, they could emulate the double-inhibition mecha- nisms of the basal ganglia (Gurney et al., 2001), dynamically tuned selection pro cesses, random ex- ploratory selections, and fo cused ‘lo cking-in’ selec- tions (e.g., see Sc hroll and Hamk er, 2013; Prescott et al., 2006; Fiore et al., 2014). At the same time, cortical winner-take-all pro cesses could contribute to tune selections at finer levels (e.g., Mysore and Kothari, 2020; Arb er and Costa, 2022). A t last, lo c k-in mechanisms could support the prolonged ac- tiv ation of specific sub-GINPs (e.g., a distal goal during planning; Baldassarre et al., 2013). 6.1.4. Motivation c omp onent: guiding the manipu- lation of GINPs The motiv ation comp onent should guide the ma- nipulator op erations on the sub-GINPs b oth di- rectly and indirectly via goals. Low-lev el moti- v ations (e.g., extrinsic motiv ations) could directly bias the op erations of the manipulator sub-GINPs. In addition, motiv ations could guide the forma- tion/activ ation of goal representations, in turn guiding the manipulator to p erform goal-directed manipulations (e.g., during planning or problem solving; Santucci et al., 2016; Rueck ert et al., 2016; Basanisi et al., 2020; Baldassarre et al., 2013). Mo- tiv ations could also bias the acquisition of task- directed represen tations and not only guide their selection (Granato et al., 2021). Differen t types of motiv ations could pla y differ- en t roles. Extrinsic motiv ations could b e imple- men ted with different mechanisms assigning v alence to stimuli and other cognitive con tents based on primary (innate) v alues related to the acquisition of material resources (Ty e, 2018). These v alence should bias b oth selection and learning pro cesses. On this basis, P avlo vian asso ciativ e learning mech- anisms could assign a se c ondary v alence to previ- ously neutral stim uli (as done in Mannella et al., 2016; Mattera et al., 2020). So cial motiv ations could w ork on the basis of similar mechanisms but rely on social stimuli having a primary v alence (e.g., see Alfieri et al., 2022). In trinsic motiv ations w ould require different mec hanisms where the primary-v alence stimuli orig- inate in the system itself when it acquires kno wl- edge and skills (Baldassarre, 2011; Baldassarre and Mirolli, 2013). No velt y could b e supp orted b y pattern recognition mechanisms while surprise b y mechanisms based on predictors (Barto et al., 2013), similarly to what migh t happ ens in the hip- p ocampus (Kumaran and Maguire, 2007). Com- p etence mec hanisms could rely on ‘goal-matching pro cesses’ that compare the pursued goal with the ac hieved world states (Baldassarre et al., 2013. Emotions ha ve more rarely been the sub ject of computational mo dels (Marsella et al., 2010). Mo d- els of emotional ‘appraisal’ could be for example b e used to ev aluate the outcomes of internal sim- ulations happ ening within the architecture (P aiv a et al., 2012). 6.2. T owar ds AI systems and r ob otic ar chite ctur es inspir e d by the GARIM the ory This section illustrates the indications that the GARIM theory can provide to enhance the auton- om y and effectiveness of AI and rob otic systems. 6.2.1. A daptive functions of c onscious and go al- oriente d states for AI and r ob otic systems The introduction of consciousness-like and goal- orien ted pro cesses into AI and robotic architectures could contribute to enhance several asp ects of them. The follo wing paragraphs consider the ma jor limi- tations of the current AI and rob otic systems, sho w- ing how mechanisms and functions inspired by the GARIM theory might contribute to face them. Flexibility. Flexibility is still a relev ant limitation of current AI systems. In particular, they are usu- ally incapable of coping with new tasks or new con- ditions and to solve problems with partial knowl- edge (Hassabis et al., 2017; Lak e et al., 2017; Mar- cus and Davis, 2019), although things migh t b e c hanging with the most recent Large Language Mo dels (LLM) discussed b elow. The GARIM the- ory prop oses that human behaviour flexibilit y de- p ends on the brain capacit y to in ternally manip- ulate the represen tations of goal-relev an t elements (e.g., ob jects, goals, actions). These manipulations giv e humans the ability to actively adjust and inte- grate the knowledge gained in previous e xperiences to cop e with nov el goals and conditions and to im- pro ve the alignmen t of behaviour to goals and of these to ultimate v alues. Therefore, the integration of mec hanisms inspired by the GARIM theory could b oost the flexibilit y of AI and robotic architectures. 27 L e arning sp e e d. The learning efficiency is a second ma jor limitation of curren t AI and rob otic systems. In particular, they are time consuming and need v ery large datasets to learn (Lake et al., 2017; Mar- cus and Da vis, 2019; Ullman, 2019). The GARIM theory introduces the sup er-ordinate representation manipulation function called Conscious Know le dge T r ansfer (CKT). Based on the four GARIM op- erations (abstraction, sp ecification, decomp osition, comp osition), CKT should make the system able to transfer knowledge b etw een tasks and domains. In particular, the introduction of this function into AI and rob otic arc hitectures could accelerate their learning pro cesses. In addition, it might allow the solution of tasks with few or no direct exp erience on them (‘few/zero shot learning’, Pourpanah et al., 2022). Cr e ativity. Creativit y and imagination are strongly limited in AI/rob otic systems (Hassabis et al., 2017; Lak e et al., 2017; Marcus and Davis, 2019). The GARIM theory p ostulates that goal-directed top- do wn manipulations of perceptual and w orking- memory representations lead to generative and cre- ativ e pro cesses. The developmen t of AI/robotic arc hitectures with these manipulation functions should b o ost their skills, for example making them able to elab orate creativ e solutions for problems. Human-AI value alignment. Man y authors argue that AI systems should b e able to in teract safely with humans, aligning their v alues and goals with ours (Harari, 2016; Bostrom, 2014; Gabriel, 2020). The GARIM theory pro vides some suggestions on ho w this could b e done. First, new architectures based on the GARIM theory would b e more flexible, th us facilitating interactions with humans. Mre- o ver, they w ould hav e a motiv ation component, th us facilitating the design of human-lik e v alue sys- tems (Dignum, 2018). In addition, they would b e able to consider emotional issues, an imp ortan t ele- men t to hav e appropriate interactions with h umans (Huang et al., 2019). Finally , the very function of consciousness proposed by the GARIM theory (alignmen t of o wn actions, goals and v alues through the manipulation of internal represen tations) migh t pro vide AI systems/robotic architectures with the fundamen tal cognitiv e abilities to align with human v alues (Brian, 2020). 6.2.2. Co gnitive R ob otics and Machine Conscious- ness: designing AI and R ob otics systems on the b asis of the GARIM the ory This section giv es initial indications on how im- plemen ting functions inspired b y the GARIM the- ory in curren t AI/robotic algorithms and architec- tures. Figure 8 illustrates a general scheme that migh t b e follo w ed to design specific AI and robotics systems based on the GARIM theory . Per c eptual c omp onent. This comp onen t should im- plemen t abstraction and generativity mechanisms. Regarding abstraction, ‘conv olutional neural net- w orks’ (CNNs; Goo dfellow et al., 2017) and ‘deep b elief netw orks’ (DBNs; Hinton et al., 2006; Hin- ton, 2002) are suitable approaches. Indeed, they can learn ‘features’ of input patterns at multi- ple lev els of abstraction. Regarding generativity , three relev ant ‘families’ of mo dels ha ve b een pro- p osed (Go odfellow et al., 2017): DBNs , considered ab o v e, ‘v ariational auto encoders’ (V AEs; Kingma and W elling, 2013), and ‘generativ e adv ersarial net- w orks’ (GANs; Go odfellow et al., 2014). Although these mo dels can b e exploited to solv e sev eral tasks, they still show limitations that might prev ent their use to implement manipulativ e func- tions prop osed by the GARIM theory . CNNs are not generative and are trained with a sup ervised algorithm. This feature makes these net works less useful for autonomous agents. V AEs are based on t wo distinct com ponents, a bottom-up abstraction comp onen t (‘enco der’) and a top-do wn generative comp onen t (‘deco der’). As a consequence, they cannot easily integrate manipulativ e processes b e- cause they would require t wo distinct manipula- tors. GANs are formed by a ‘discriminator com- p onen t’ and a ‘generator comp onen t’: the latter could b e useful to implemen t manipulativ e func- tionalities, while the former could b e used to dis- tinguish b et ween imagined and p erceiv ed stimuli. Unfortunately , the ‘generative sto c hastic engine’ of b oth V AEs and GANs is limited. In particular, it is lo cated in the latent space of V AEs and in the discriminator of GANs. Therefore, the tw o net- w orks cannot ha ve generativity at multiple levels of abstraction. In terestingly , DBNs show a bidi- rectional arc hitecture implementing b oth bottom- up abstraction and top-down generative processes. Moreo ver, they show t wo in teresting features: (a) their ‘generative engine’ is distributed into all its sto c hastic-units, thus supp orting generativity at m ultiple lev els of abstraction and (b) their unsup er- 28 Figure 8: Blueprint of an AI architecture based on the GARIM theory . The figure shows some AI algorithms that could be used to implemen t the functions of the GARIM theory components. Bold text: names of the components; Plain text: functions; Italics text, in brack ets: algorithms/mo dels; Dash-highligh ted text: representations on whic h consciousness pro cesses operate. vised learning mechanisms can b e in tegrated with reinforcemen t learning mechanisms, thus balancing represen tational requirements and task demands (Granato et al., 2022b). Working memory c omp onent. This component should supp ort the information reverberation in the absence of the corresp onding patterns from sensors or internal processes. The comp onen t should be able to learn whic h patterns to store and whic h not, also on the basis of goals. RNNs, in tro duced abov e, are a first pow erful tool usable to implement working memory . This capac- it y is based on an arc hitecture having re-en trant connections and thus capable of dynamically stor- ing information (Choi et al., 2018). L ong-short term memories (LSTM; Ho c hreiter and Sc hmid- h ub er, 1997) are netw orks based on units with a ‘gated self-connection’ and gates in input and out- put connections. The op ening/closing of the gates can upload/do wnload information in the unit, mak- ing it capable of storing memories for long times. These net works are commonly used to s olv e classi- fication and regression tasks with input sequences. Ho wev er, they hav e recen tly b een up dated with ad- ditional mechanisms that can supp ort delib erativ e (goal-directed) pro cesses as needed b y the GARIM theory (e.g., see Jung et al., 2019b). Neur al T uring machines (Grav es et al., 2014; W a yne et al., 2018) are neural netw orks that supp ort delib erativ e pro- cesses. These netw orks use ‘w orking memory slots’ that are based on numerical vectors. These slots are read/written b y ‘neural heads’ that are trainable with gradient-based algorithms. These features al- lo w these netw orks to implement trainable logic-like reasoning. How ev er, the pre-defined lev el of ab- straction of these memory slots make them unsuit- able to implement the GARIM op erations of com- p osition/decomposition, thus limiting their flexibil- it y . Manipulator c omp onent. This comp onen t should implemen t tw o main functions. First, it should supp ort the autonomous learning and performance of the goal-directed manipulation of representations (states, goals, actions, etc.). Second, it should sup- p ort the goal-directed adaptation/tuning of these manipulation pro cesses. A num b er of AI mech- anisms, introduced ab o ve, can b e used to imple- men t working memories and ‘neural heads’, or other mec hanisms, to ‘read/write’ suc h memories. These mec hanisms can b e imp ortan t means to implement the manipulation of representations. The implementation of goal-directed processes 29 also requires the p erformance of a n umber of structured and temp orised op erations, such as the goals/sub-goals activ ation/de-activ ation. Exam- ples of these are: the generation and search of cor- rect action sequences, the prediction of actions out- comes, the exc hange of information b et ween the differen t comp onen ts of the system. These op era- tions are relatively easy to implement with symbolic represen tations and programming con trols (e.g., ‘if- then’ and ‘lo op’ op erations; Russell and Norvig, 2016) but very difficult to implement with neu- ral mec hanisms. Curren t systems th us tend to b e based on h ybrid neural/symbolic mec hanisms. This is an imp ortan t op en problem as the non-neural parts of the mo dels could obstacle the information in tegration capabilities of the system. Hybrid sys- tems (Sun, 2016; Konidaris et al., 2018; Oddi et al., 2019) implemen t low-lev el cognitive pro cesses based on neural representations and learning algorithms. A t the same time, they implement high-level cog- nitiv e pro cesses based on symbolic representations. This double representation format allo ws them, for example, to implement sym b olic PDDL planning while using neural mechanisms to implement sen- sorimotor pro cesses. These approac hes hav e lim- itations for our scope. In particular, they intro- duce inhomogeneous represen tations at the lo w and high represen tation levels, requiring differen t mec h- anisms to manipulate them. Neur al T uring ma- chines and models lik e MERLIN (Grav es et al., 2014; W ayne et al., 2018) use memory slots and neu- ral heads to perform complex tasks that require the ac hievemen t of m ultiple subgoals. This approac h is mainly used to solve single reactiv e tasks but it can also b e used to solve delib erativ e problems (Chaplot et al., 2021). Neur osymb olic AI (for a review see Garcez and Lamb, 2020), and in par- ticular recent visual planning systems (Jung et al., 2019b; Nair et al., 2018), p erform planning task on the basis of goal-directed pro cesses and distributed represen tations (states, goals, actions, etc.). These pro cesses allow high flexibility , supp orting gener- alisation capabilities that cannot b e ac hieved by sym b olic planning/problem solving. Ho wev er, for no w they cannot comp ose/decompose the manipu- lated elements. T r ansformers (V asw ani et al., 2017) implemen t neural internal atten tion mechanisms and dynamic circuits. Their memory and atten- tion units are in tegrated within the trainable input- output lay ers of neural netw orks. These mo dels are v ery effective in recalling any learnt or acquired in- formation, even if exp erienced muc h earlier. T rans- formers hav e b een mainly used to successfully solve natural language pro cessing tasks but require mas- siv e sup ervised-learning training (Blakeman and Maresc hal, 2022). These systems hav e the p oten- tial to also supp ort delib erativ e pro cesses (Chaplot et al., 2021) and to b e h ybridised with sensorimo- tor components (Driess et al., 2023). In addition, they hav e b een indicated as relev an t to implement consciousness-lik e pro cesses (Bengio, 2017). How- ev er, the functioning of AI systems based on trans- formers is still p oorly understo o d (Ag ¨ uera y Arcas, 2022; Sriv asta v a and al., 2022). In this resp ect, in the future it might be interesting to ev aluate if ma- nipulation op erations similar to those prop osed by the GARIM theory actually take place within AI systems based on transformers. Motivation c omp onent. Extrinsic motiv ations are usually emulated trough reward signals (Sutton and Barto, 1998). Moreov er, ‘pseudo-rewards’ can b e used to guide mo del-based hierarc hical rein- forcemen t learning based on goal-matching ev ents (Botvinic k et al., 2008). In trinsic motiv ations hav e demonstrated to effec- tiv ely supp ort the autonomous acquisition of knowl- edge of rob ots (Baldassarre and Mirolli, 2013). In- deed, in trinsic motiv ation mechanisms can driv e the in vestigation and learning of nov el/surprising ex- p eriences, leading to the acquisition of new state represen tations and mo dels (Sc hmidh ub er, 1991; Oudey er et al., 2007; Barto et al., 2013; Cartoni and Baldassarre, 2018). Moreo ver, they can lead to the acquisition of ‘intrinsic goals’ (autonomously found) and motor skills to accomplish them (Barto et al., 2004; Santucci et al., 2016; Nair et al., 2018). In trinsic motiv ations are commonly used to guide in telligent machines and rob ots to seek kno wledge in the external environmen t. Instead, according to the GARIM prop osal they could guide the internal building of the knowledge that the agen t lacks. There are few AI approaches that emulate the generation of emotions (for reviews see Paiv a et al., 2012; Mirolli et al., 2010; Sun et al., 2016). These mo dels could b e used as a starting p oin t for imple- men ting emotion-based ev aluation of internal rep- resen tations. Op en chal lenges: what is missing?. The t wo ar- c hitecture schemes w e prop osed for guiding the dev elopment of computational mo dels (Figure 7) and AI/rob otic architectures (Figure 8) include the main elements that should supp ort conscious 30 and flexible artificial systems. F or example, they include the main features of machine conscious- ness systems (self-mo delling, information broad- casting, higher-lev el representations, attention pro- cesses, and information integration; Reggia, 2013). Moreo ver, they include the fundamentals axioms of Mac hine Consciousness (world mo dels, imagina- tion, attention, planning, and affective ev aluation; Aleksander, 1995) . How ev er, critical elements for building conscious machines may still b e missing. First, the four macro-systems prop osed by the GARIM theory require important low-lev el func- tions to supp ort the emergence of GINPs. F or ex- ample, the brain sho ws a high capacit y for gen- erating asso ciations and av oiding un b ounded ac- tiv ations. These capacities are based on grid-like circuits and finely regulated inhibitory pro cesses. These features are missing in common artificial neu- ral net w ork arc hitectures, whic h fa vour b ottom- up/top-do wn directional information flo ws with few recurrences (Lynn and Bassett, 2019). Second, the brain exhibits highly dynamic pro cesses that could b e based on fixed-p oin t/cycle/strange attractors. ANNs are still not able to fully emulate these pro- cesses. These elemen ts migh t b e needed to imple- men t the GARIM op erations on sub-GINPs (Break- sp ear, 2017). Third, the flexible selection func- tions implemen ted b y the basal ganglia-thalamo- cortical lo ops are only partially captured by cur- ren t neural systems. F ourth, strongly-coupled sen- sorimotor lo ops engaged by animals with the en- vironmen t are often absen t in AI systems. More- o ver, curren t rob ots hav e still a very limited au- tonom y to interact with the en vironment. Agen t- en vironment in teractions might instead b e v ery im- p ortan t to acquire in ternal representations strongly coupled with the real en vironment. Last, till re- cen tly AI/robotic systems lack ed the capacit y to suitably in tegrate language with sensorimotor ex- p erience, in particular they lack ed a meaning and understanding grounded on sensorimotor experi- ence. Recently , how ever, large language mo dels ha ve b een argued to acquire some meaning ev en without sensorimotor grounding (Y un et al., 2021; Ab dou et al., 2021; Ag ¨ uera y Arcas, 2022; Sriv as- ta v a and al., 2022), and grounding might b e re- alised very so on with systems integrating language and sensorimotor capabilities (Driess et al., 2023). The fact that these mo dels are based on transform- ers, whic h migh t p erform op erations very similar to the representation manipulations p ostulated b y the GARIM theory , hence represents an in teresting topic for future inv estigations. Ov erall, the realisation and integration of all these elements is still a great op en challenge. Much of the flexibility of the brain is based on its highly structured and integrated architecture, which seems difficult to repro duce in artificial systems. Indeed, the brain in tegrates habitual and goal-directed pro- cesses and it is the pro duct of a long ev olution- ary pro cess that is hard to repro duce in machines (Baldassarre et al., 2017; Baldassarre and Granato, 2020; Caligiore et al., 2019; Ullman, 2019). Produc- ing conscious intelligen t machines without relying on such a highly in tegrated arc hitecture is there- fore a great challenge. 7. Conclusions In this w ork w e in tro duce the Go al-Aligning R ep- r esentation Internal Manipulation (GARIM) the- ory of flexible goal-directed cognition and con- sciousness. The cen tral idea of the GARIM the- ory is that conscious states supp ort the active manipulation of internal representations, making them more aligned with the goals pursued. This goal-orien ted alignment leads to the generation of the necessary knowledge to face no vel situa- tions and goals and to make goal-directed b e- ha viour more flexible and effectiv e. The GARIM theory p ostulates that a conscious goal-directed b eha viour is characterised by five distinctive el- emen ts. First, consciousness serves the adapta- tion of goal-directed behaviours. In particular, consciousness pro cesses supp ort the goal-aligning manipulation of internal represen tations, in turn b oosting the flexibilit y of goal-directed behaviour. Second, the theory hypotheses the existence of ‘Goal-based Integrated Neural Patterns’ (GINPs). These are distributed active neural represent ations that (a) are consciously p erceiv ed and thus in- ten tionally manipulable (consciousness level), and (b) are closely related the pursued goals (goal- relev ance). Different levels of these t w o dimen- sions lead to representations c haracterised by dif- feren t lev els of consciousness (Non-GINPs, Pre- GINPs, T emp-GINPs, GINPs) and representational proprieties (e.g., information integration). Third, the GARIM theory sp ecifies that goal-directed ma- nipulations rely on four key ‘comp onen ts’, namely four partially o v erlapping anatomo-functional brain macro systems (perceptual w orking-memory , ab- stract working-memory , internal manipulator, mo- tiv ational component). F ourth, previous systems 31 giv e rise to four classes of computational op era- tions (GARIM op erations) that supp ort the repre- sen tation manipulations (abstraction, sp ecification, decomp osition, comp osition). Fifth and last, the GARIM theory introduces the concept of ‘GARIM agency’, a sense of agency that emerges from con- scious goal-directed processes and, in particular, represen tation manipulations. These manipula- tions lead to the generation of a sub jective inter- nal realit y supp orted by three key features: self- mo del, emotional/p erceptual vividness, and manip- ulation con trol. On the basis of the GARIM agency and its key features, the theory proposes different lev els of conscious states (from reactive b eha viours to highly flexible goal-directed b eha viours). In ad- diction to clarifying neuro-computational pro cesses at the basis of conscious and flexible goal-directed b eha viours, the GARIM theory has b oth scien- tific and tec hnological implications. F or example, it clarifies some asp ects of sub jective exp erience and agency , also introducing a p oten tial quanti- tativ e scale. Moreo ver, it accounts for several el- emen ts of curren t theories of consciousness, in te- grating them into a common functional and com- putational framework that fo cuses on goal-directed pro cesses. The GARIM theory also generates in- sigh ts for exp erimen tal and clinical fields. In par- ticular, it prop oses clinical insights, experimental predictions and new ideas for building experimental proto col of goal-directed behaviour and conscious- ness. A t last, the GARIM theory furnishes indica- tions for building new computational mo dels, and AI/rob otic architectures. In particular, it prop oses that conscious goal-aligning manipulations of rep- resen tations could enable AI/robotic architectures to achiev e human-lik e flexibility and general in tel- ligence. 8. Ac kno wledgements This work has received funding from the Eu- rop ean Union’s Horizon 2020 Research and In- no v ation Program with the pro jects ‘GO AL- Rob ots – Goal-based Op en-ended Autonomous Learning Rob ots’, GA N. 713010, ‘HBP – Hu- man Brain Pro ject SGA3’, GA N. 945539; and from the Horizon Europ e Program with pro jects ‘PILLAR-Rob ots - Purp oseful Intrinsically moti- v ated Lifelong Learning Autonomous Rob ots’, GA N. 101070381, and ‘EBRAINS-Italy - Europ ean Brain ReseArch INfrastructureS Italy’, PNRR N. IR000001, CUP B51E22000150006. W e thank Emilio Cartoni and Andrea Mattera for the useful feedbac k on the early versions of the manuscript. References Aaronson, S., 2014. Why i am not an in tegrated information theorist (or, the unconscious expander). Shtetl Optimized: The Blog of Scott Aaronson . Abdou, M., Kulmizev, A., Hershcovich, D., F rank, S., Pa vlic k, E., Søgaard, A., 2021. Can Language Mo dels En- code Perceptual Structure Without Grounding? A Case Study in Color. T echnical Rep ort. arXiv. doi: 10.48550/ arXiv.2109.06129 , arXiv:2109.06129 . Aleksander, I., 1995. Artificial neuro consciousness an up- date, in: International W orkshop on Artificial Neural Net- works, Springer. pp. 566–583. Aleksander, I., Dunmall, B., 2003. Axioms and tests for the presence of minimal consciousness in agen ts. Journal of Consciousness Studies 10, 7–18. Alfieri, V., Mattera, A., Baldassarre, G., 2022. Neural cir- cuits underlying social fear in ro den ts: An integrativ e computational model. F rontiers in systems neuroscience 16, 841085. doi: 10.3389/fnsys.2022.841085 . Amaral, D.G., 2002. The primate amygdala and the neuro- biology of so cial behavior: implications for understanding social anxiety . Biol Psychiatry 51, 11–17. Anderson, M.L., 2003. Embo died cognition: A field guide. Artificial intelligence 149, 91–130. Arber, S., Costa, R.M., 2022. Net working brainstem and basal ganglia circuits for mo vemen t. Nature reviews Neu- roscience 23, 342–360. doi: 10.1038/s41583- 022- 00581- w . Ag¨ uera y Arcas, B., 2022. Do large language models under- stand us? Daedalus 151, 183–197. Aru, J., Bachmann, T., Singer, W., Melloni, L., 2012. Dis- tilling the neural correlates of consciousness. Neuroscience & Biobehavioral Reviews 36, 737–746. Atherton, M., Zhuang, J., Bart, W.M., Hu, X., He, S., 2003. A functional mri study of high-level cognition. i. the game of chess. Cognitive Brain Researc h 16, 26–31. Baars, B.J., 1997. In the theatre of consciousness. global workspace theory , a rigorous scientific theory of conscious- ness. Journal of Consciousness Studies 4, 292–309. Baars, B.J., 2005. Global w orkspace theory of conscious- ness: tow ard a cognitive neuroscience of human exp eri- ence. Progress in brain research 150, 45–53. Baars, B.J., F ranklin, S., Ramso y , T.Z., 2013. Global workspace dynamics: cortical ”binding and propagation” enables conscious conten ts. F rontiers in psyc hology 4. Baars, B.J., Ramsøy , T.Z., Laureys, S., 2003. Brain, con- scious exp erience and the observing self. T rends in neu- rosciences 26, 671–675. Baldassarre, G., 2011. What are in trinsic motiv ations? a bi- ological p ersp ectiv e, in: Cangelosi, A., T riesch, J., F asel, I., Rohlfing, K., Nori, F., Oudey er, P .Y., Sc hlesinger, M., Nagai, Y. (Eds.), Pro ceedings of the International Conference on Dev elopment and Learning and Epigenetic Robotics (ICDL-EpiRob-2011), F rankfurt Institute of Ad- v anced Studies (FIAS). IEEE, New Y ork, NY. pp. e1–8. F rankfurt am Main, Germany , 24–27/08/11. Baldassarre, G., Granato, G., 2020. Goal-directed manip- ulation of in ternal representations is the core of general- domain intelligence. Journal of Artificial General Intelli- gence 11, 19–23. 32 Baldassarre, G., Mannella, F., Fiore, V.G., Redgrav e, P ., Gurney , K., Mirolli, M., 2013. Intrinsically motiv ated action-outcome learning and goal-based action recall: A system-level bio-constrained computational mo del. Neu- ral Netw orks 41, 168–187. Baldassarre, G., Mannella, F., Santucci, V.G., Somogyi, E., Jacquey , L., Hamilton, M., O’Regan, J.K., 2018. Action- outcome con tingencies as the engine of op en-ended learn- ing: computational mo dels and developmen tal exp eri- ments, in: The 8th IEEE International Conference on De- velopmen t and Learning and Epigenetic Rob otics (ICDL- EpiRob2018), pp. E1–8. T okyo, Japan, 16–20 September 2018. Baldassarre, G., Mirolli, M. (Eds.), 2013. Intrinsically moti- v ated learning in natural and artificial systems. Springer, Berlin. Cost 91.62 euros, pp. 458, 82 illustrations, 55 il- lustrations in color. Baldassarre, G., San tucci, V.G., Cartoni, E., Caligiore, D., 2017. The architecture challenge: F uture artificial- intelligence systems will require sophisticated architec- tures, and knowledge of the brain might guide their construction. Behavioral and Brain Sciences 40, e254. doi: 10.1017/s0140525x17000036 . Balleine, B.W., Dickinson, A., 1998. Goal-directed instru- mental action: contingency and incentiv e learning and their cortical substrates. Neuropharmacology 37, 407– 419. Balleine, B.W., Ostlund, S.B., 2007. Still at the c hoice-p oin t: action selection and initiation in instrumental condition- ing. Ann N Y Acad Sci 1104, 147–171. doi: 10.1196/ annals.1390.006 . Barak, O., Tso dyks, M., 2014. Working mo dels of working memory. Current Opinion in Neurobiology 25, 20–24. Barto, A.G., Mirolli, M., Baldassarre, G., 2013. Nov elty or surprise? F rontiers in Psychology – Cognitive Science 4, e1–15. Edited by: T om Stafford, University of Sheffield, UK Reviewed by: Karl F riston, Universit y College Lon- don, UK; Nathan F.Lep ora, The Universit y of Sheffield, UK. Barto, A.G., Singh, S., Chentanez, N., 2004. Intrinsically motiv ated learning of hierarchical collections of skills, in: T riesch, J., Jebara, T. (Eds.), International Conference on Developmen tal Learning (ICDL2004), New Y ork, NY. pp. 112–119. Basanisi, R., Brov elli, A., Cartoni, E., Baldassarre, G., 2020. A spiking neural-netw ork mo del of goal-directed behaviour. Plos Computational Biology . Bechara, A., Damasio, A.R., 2005. The somatic marker hy- pothesis: A neural theory of economic decision. Games and economic behavior 52, 336–372. Bengio, Y., 2017. The consciousness prior. arXiv preprint arXiv:1709.08568 . Benson, N., Hulac, D.M., Kranzler, J.H., 2010. Inde- pendent examination of the wec hsler adult intelligence scale—fourth edition (w ais-iv): what does the wais-iv measure? Psychological Assessment 22, 121. Berg, E.A., 1948. A simple ob jective technique for measuring flexibility in thinking. The Journal of general psychology 39, 15–22. Berke, J.D., 2018. What do es dopamine mean? Nature neuroscience 21, 787–793. Blakeman, S., Mareschal, D., 2022. Selective particle at- tention: Rapidly and flexibly selecting features for deep reinforcement learning. Neural Net works 150, 408–421. Borghi, A.M., Cimatti, F., 2010. Embodied cognition and beyond: acting and sensing the b ody . Neuropsyc hologia 48, 763–773. Bortolon, C., Macgregor, A., Cap devielle, D., Raffard, S., 2018. Apath y in schizophrenia: A review of neuropsyc ho- logical and neuroanatomical studies. Neuropsychologia 118, 22–33. Bostrom, N., 2014. Sup erin telligence: P aths, Dangers, Strategies. Oxford Universit y Press, Oxford. Botvinick, M.M., Niv, Y., Barto, A., 2008. Hierarchi- cally organized b eha vior and its neural foundations: A reinforcement-learning p erspective. Cognition 113, 262– 280. Brav er, T.S., Bongiolatti, S.R., 2002. The role of frontopo- lar cortex in subgoal pro cessing during working memory . Neuroimage 15, 523–536. Breakspear, M., 2017. Dynamic mo dels of large-scale brain activity. Nature Neuroscience 20, 340–352. Brian, C., 2020. The Alignmen t Problem: Machine Learning and Human V alues. W.W. Norton & Company , New Y ork. Brown, R., Lau, H., LeDoux, J.E., 2019. Understanding the higher-order approach to consciousness. T rends in cognitive sciences 23, 754–768. Bubeck, S., Chandrasek aran, V., Eldan, R., Gehrke, J., Horvitz, E., Kamar, E., Lee, P ., Lee, Y.T., Li, Y., Lund- berg, S., Nori, H., Palangi, H., Ribeiro, M.T., Zhang, Y., 2023. Sparks of artificial general intelligence: Early ex- periments with gpt-4. doi: 10.48550/ARXIV.2303.12712 . Burk art, J.M., Sch ubiger, M.N., v an Sc haik, C.P ., 2017. The evolution of general intelligence. Behavioral and Brain Sciences 40. Cabanac, M., 2002. What is emotion? Behavioural pro cesses 60, 69–83. Caligiore, D., Arbib, M.A., Miall, C.R., Baldassarre, G., 2019. The sup er-learning hypothesis: Integrating learn- ing pro cesses across cortex, cereb ellum and basal ganglia. Neuroscience and Biobehavioral Reviews 100, 19–34. Carter, O., Hohwy , J., V an Boxtel, J., Lamme, V., Blo c k, N., Ko c h, C., Tsuchiy a, N., 2018. Conscious machines: Defining questions. Science 359, 400–400. Cartoni, E., Baldassarre, G., 2018. Autonomous discov ery of the goal space to learn a parameterized skill. Preprint arXiv 1805.07547v1. arXiv. 19 May 2018. Cerullo, M.A., 2015. The Problem with Phi: A Critique of Integrated Information Theory. PLoS Comput Biol 11, e1004286. Chalmers, D.J., 1995. F acing up to the problem of conscious- ness. Journal of consciousness studies 2, 200–219. Chaplot, D.S., P athak, D., Malik, J., 2021. Differentiable Spatial Planning using T ransformers. ArXiv Preprint abs/2112.01010. Chelazzi, L., Perlato, A., Santandrea, E., Della Lib era, C., 2013. Rewards teach visual selectiv e attention. Vision research 85, 58–72. Chiappe, D., MacDonald, K., 2005. The evolution of domain-general mec hanisms in intelligence and learning. The Journal of general psychology 132, 5–40. Choi, M., Matsumoto, T., Jung, M., T ani, J., 2018. Gener- ating Goal-Directed Visuomotor Plans Based on Learning Using a Predictiv e Co ding-t yp e Deep Visuomotor Recur- rent Neural Network Mo del. arXiv abs/1803.02578v2. Christoff, K., Irving, Z.C., F ox, K.C., Spreng, R.N., Andrews-Hanna, J.R., 2016. Mind-wandering as sp on- taneous thought: a dynamic framework. Nature Reviews Neuroscience 17, 718–731. Chrysikou, E.G., Mot yk a, K., Nigro, C., Y ang, S.I., 33 Thompson-Schill, S.L., 2016. F unctional fixedness in cre- ative thinking tasks dep ends on stim ulus mo dality . Psy- chology of Aesthetics, Creativity , and the Arts 10, 425. Clark, A., 2013. Whatever next? predictive brains, situated agents, and the future of cognitive science. Beha vioral and brain sciences 36, 181–204. Cleeremans, A., 2007. Consciousness: the radical plasticity thesis. Progress in brain research 168, 19–33. Cleeremans, A., 2011. The radical plasticit y thesis: how the brain learns to b e conscious. F rontiers in psychology 2, 86. Cleeremans, A., Ac houi, D., Beauny , A., Keuninc kx, L., Martin, J.R., Mu ˜ noz-Moldes, S., V uillaume, L., De Heer- ing, A., 2020. Learning to b e conscious. T rends in cogni- tive sciences 24, 112–123. Cow ey , A., Sto erig, P ., 1995. Blindsight in monkeys. Nature 373, 247–249. Damasio, A.R., 1989. The brain binds entities and even ts by multiregional activ ation from conv ergence zones. Neural Computation 1, 123–132. Damasio, A.R., 1998. Emotion in the p ersp ectiv e of an inte- grated nerv ous system. Brain research reviews 26, 83–86. Damasio, A.R., Meyer, K., 2009a. Consciousness: An ov erview of the phenomenon and of its p ossible neural basis. The neurology of consciousness: Cognitive neuro- science and neuropathology , 3–14. Damasio, A.R., Meyer, K., 2009b. Consciousness: An ov erview of the phenomenon and of its p ossible neural basis. The neurology of consciousness: Cognitive neuro- science and neuropathology , 3–14. Day an, P ., Abb ott, L.F., 2001. Theoretical Neuroscience. The MIT Press, Cambridge, MA. Dehaene, S., Changeux, J.P ., 2005. Ongoing sp on taneous activity controls access to consciousness: a neuronal model for inatten tional blindness. PLoS biology 3, e141. doi: 10.1371/journal.pbio.0030141 . Dehaene, S., Changeux, J.P ., 2011. Experimental and the- oretical approaches to conscious pro cessing. Neuron 70, 200–227. Dehaene, S., Changeux, J.P ., Naccac he, L., 2011. The global neuronal w orkspace mo del of conscious access: from neu- ronal architectures to clinical applications, in: Char- acterizing consciousness: from cognition to the clinic?. Springer, pp. 55–84. Dehaene, S., Kerszb erg, M., Changeux, J.P ., 1998a. A neu- ronal mo del of a global workspace in effortful cognitiv e tasks. Proceedings of the National Academy of Sciences of the United States of America 95, 14529–14534. Dehaene, S., Kerszb erg, M., Changeux, J.P ., 1998b. A neu- ronal mo del of a global workspace in effortful cognitiv e tasks. Pro ceedings of the national Academ y of Sciences 95, 14529–14534. Dehaene, S., Lau, H., Kouider, S., 2017. What is conscious- ness, and could machines hav e it? Science (New Y ork, N.Y.) 358, 486–492. doi: 10.1126/science.aan8871 . Dehaene, S., Naccac he, L., 2001. T o wards a cognitive neuro- science of consciousness: basic evidence and a workspace framework. Cognition 79, 1–37. Dehaene, S., Sergent, C., Changeux, J.P ., 2003. A neuronal netw ork model linking sub jectiv e rep orts and ob jective physiological data during conscious p erception. Proceed- ings of the National Academ y of Sciences 100, 8520–8525. Del Pin, S.H., Sk´ ora, Z., Sandberg, K., Overgaard, M., Wierzcho´ n, M., 2021. Comparing theories of conscious- ness: why it matters and how to do it. Neuroscience of Consciousness 2021, niab019. Delatour, B., Gisquet-V errier, P ., 2000. F unctional role of rat prelimbic-infralim bic cortices in spatial memory: evi- dence for their inv olvemen t in attention and b eha vioural flexibility . Beha vioural brain research 109, 113–128. Diamond, A., 2013. Executiv e functions. Annu Rev Psyc hol 64, 135–168. Diehl, P .U., Co ok, M., 2015. Unsup ervised learning of digit recognition using spike-timing-dependent plasticit y . F ron- tiers in Computational Neuroscience 9, 99. doi: 10.3389/ fncom.2015.00099 . Dignum, V., 2018. Ethics in artificial intelligence: introduc- tion to the special issue. Ethics and Information T ec hnol- ogy 20, 1–3. Doerig, A., Sch urger, A., Herzog, M.H., 2021. Hard criteria for empirical theories of consciousness. Cognitiv e neuro- science 12, 41–62. Donnarumma, F., Costantini, M., Ambrosini, E., F riston, K., Pezzulo, G., 2017. Action perception as hypothesis testing. Cortex 89, 45–60. Driess, D., Xia, F., Sa jjadi, M.S.M., Lynch, C., Chowdh- ery , A., Ich ter, B., W ahid, A., T ompson, J., V uong, Q., Y u, T., Huang, W., Cheb otar, Y., Sermanet, P ., Duc k- worth, D., Levine, S., V anhouc ke, V., Hausman, K., T ou- ssaint, M., Greff, K., Zeng, A., Mordatch, I., Florence, P ., 2023. P aLM-E: An Embo died Multimo dal Language Model. T echnical Report 2303.03378. arXiv. Dunck er, K., 1945. On Problem Solving. Number 58 in Psychological Monographs, American Psyc hological As- sociation. Ekman, P ., Davidson, R.J., 1994. The Nature of emotion: fundamental questions. Oxford Universit y Press, New Y ork. Ell, S.W., Marchan t, N.L., Ivry , R.B., 2006. F o cal putamen lesions impair learning in rule-based, but not information- integration categorization tasks. Neuropsyc hologia 44, 1737–1751. Ell, S.W., W einstein, A., Ivry , R.B., 2010. Rule-based cate- gorization deficits in focal basal ganglia lesion and parkin- son’s disease patients. Neuropsychologia 48, 2974–2986. F ernandez-Duque, D., Baird, J.A., P osner, M.I., 2000. Exec- utive atten tion and metacognitive regulation. Conscious- ness and cognition 9, 288–307. F errucci, D.A., 2012. Introduction to “this is watson”. IBM Journal of Research and Development 56, 1–1. Fiore, V.G., Sp erati, V., Mannella, F., Mirolli, M., Gurney , K., Firston, K., Dolan, R.J., Baldassarre, G., 2014. Keep focussing: striatal dopamine multiple functions resolv ed in a single mechanism tested in a simulated humanoid robot. F ron tiers in Psychology 5, e1–17. doi: 10.3389/ fpsyg.2014.00124 . F ornazzari, L., F arcnik, K., Smith, I., Heasman, G.A., Ic hise, M., 1992. Violent visual hallucinations and aggression in frontal lobe dysfunction: clinical manifestations of deep orbitofrontal fo ci. The Journal of Neuropsychiatry and Clinical Neurosciences . F ox, K.C., Shi, L., Baek, S., Raccah, O., F oster, B.L., Saha, S., Margulies, D.S., Kucyi, A., P arvizi, J., 2020. Intrinsic netw ork architecture predicts the effects elicited by in- tracranial electrical stimulation of the human brain. Na- ture human b ehaviour 4, 1039–1052. F ranklin, S., Strain, S., Snaider, J., McCall, R., F aghihi, U., 2012. Global workspace theory , its lida mo del and the underlying neuroscience. Biologically Inspired Cognitive Architectures 1, 32–43. 34 F riston, K., 2018. Am i self-conscious?(or do es self- organization entail self-consciousness?). F rontiers in psy- chology 9, 579. F riston, K., FitzGerald, T., Rigoli, F., Schw arten b eck , P ., Pezzulo, G., et al., 2016. Activ e inference and learning. Neuroscience & Biobehavioral Reviews 68, 862–879. F rith, C., 1996. The role of the prefrontal cortex in self- consciousness: the case of auditory hallucinations. Philo- sophical T ransactions of the Ro yal So ciet y of London. Se- ries B: Biological Sciences 351, 1505–1512. F rith, C.D., 2007. The so cial brain? Philosophical T rans- actions of the Ro yal So ciety B: Biological Sciences 362, 671–678. F uster, J.M., 2008. The prefrontal cortex. F ourth ed., Else- vier, Oxford. F uster, J.M., Bressler, S.L., 2015. Past makes future: role of pfc in prediction. Journal of cognitiv e neuroscience 27, 639–654. Gabriel, I., 2020. Artificial intelligence, v alues, and align- ment. Minds and mac hines 30, 411–437. doi: 10.1007/ s11023- 020- 09539- 2 . Gamez, D., 2008. Progress in machine consciousness. Con- sciousness and cognition 17, 887–910. Gamez, D., 2010. Information in tegration based predictions about the conscious states of a spiking neural net work. Consciousness and cognition 19, 294–310. Gangestad, S.W., Grebe, N.M., 2017. Hormonal systems, human social b onding, and affiliation. Horm Behav 91, 122–135. Garbarini, F., Adenzato, M., 2004. At the root of em b odied cognition: cognitive science meets neuroph ysiology . Brain and cognition 56, 100–106. Garcez, A.d., Lamb, L.C., 2020. Neurosymbolic ai: the 3rd wa v e. arXiv preprint arXiv:2012.05876 . Gardner, H.E., 2000. In telligence reframed: Multiple intelli- gences for the 21st century . Hac hette UK. Gazzaley , A., Nobre, A.C., 2012. T op-down mo dulation: bridging selectiv e atten tion and working memory . T rends in cognitive sciences 16, 129–135. Goo dfellow, I., Bengio, Y., Courville, A., 2017. Deep Learn- ing. The MIT Press, Boston, MA. Goo dfellow, I., Pouget-Abadie, J., Mirza, M., Xu, B., W arde-F arley , D., Ozair, S., Courville, A., Bengio, Y., 2014. Generative adversarial nets, in: Adv ances in Neu- ral Information Pro cessing Systems, pp. 2672–2680. 10 Jun 2014. Granato, G., Baldassarre, G., 2021. Internal manipulation of p erceptual representations in human flexible cognition: A computational mo del. Neural Net works 143, 572–594. doi: 10.1016/j.neunet.2021.07.013 . Granato, G., Borghi, A.M., Baldassarre, G., 2020. A com- putational mo del of language functions in flexible goal- directed behaviour. Scientific reports 10, 1–13. Granato, G., Borghi, A.M., Mattera, A., Baldassarre, G., 2022a. A computational mo del of inner sp eech supp ort- ing flexible goal-directed behaviour in autism. Scien tific Reports 12, 14198. Granato, G., Cartoni, E., Da Rold, F., Mattera, A., Bal- dassarre, G., 2021. A Computational Mo del of Repre- sentation Learning in the Brain Cortex, Integrating Un- supervised and Reinforcement Learning. arXiv preprint, Granato, G., Cartoni, E., Da Rold, F., Mattera, A., Baldas- sarre, G., 2022b. Integrating unsupervised and reinforce- ment learning in human categorical p erception: A com- putational mo del. PLOS ONE 17, 1–32. doi: 10.1371/ journal.pone.0267838 . Granato, G., Costanzo, R., Borghi, A.M., Mattera, A., Car- ruthers, S., Rossell, S., Baldassarre, G., . Flexible cog- nition and inner-sp eec h in schizophrenia sp ectrum disor- der: Applying machine learning and mo delling in compu- tational psychiatry . Grav es, A., W a yne, G., Danihelk a, I., 2014. Neural T uring Machines. arXiv preprint 1410.5401. Gruberger, M., Ben-Simon, E., Levk ovitz, Y., Zangen, A., Hendler, T., 2011. T ow ards a neuroscience of mind- wandering. F ron tiers in human neuroscience 5, 56. Guilford, J.P ., 1967. The nature of human intelligence. McGraw-Hill. Gurney , K., Prescott, T.J., Redgrav e, P ., 2001. A compu- tational mo del of action selection in the basal ganglia. ii. analysis and simulation of b eha viour. Biological Cyb er- netics 84, 411–423. H¨ anggi, J., Br ¨ utsch, K., Siegel, A.M., J¨ anc ke, L., 2014. The architecture of the chess play er’s brain. Neuropsychologia 62, 152–162. Harari, Y.N., 2016. Homo Deus: A brief history of tomorrow. Random House. Hartley , A.A., Sp eer, N.K., 2000. Lo cating and fractionat- ing working memory using functional neuroimaging: stor- age, maintenance, and executiv e functions. Microscopy research and technique 51, 45–53. Hassabis, D., Kumaran, D., Summerfield, C., Botvinic k, M., 2017. Neuroscience-inspired artificial in telligence. Neuron 95, 245–258. doi: 10.1016/j.neuron.2017.06.011 . Hasz, B.M., Redish, A.D., 2020. Spatial enco ding in dorso- medial prefron tal cortex and hippo campus is related dur- ing delib eration. Hipp ocampus 30, 1194–1208. doi: 10. 1002/hipo.23250 . He, M., Qi, C., Lu, Y., Song, A., Hay at, S.Z., Xu, X., 2018. The sp ort expert’s attention sup eriorit y on skill-related scene dynamic by the activ ation of left medial frontal gyrus: an erp and loreta study . Neuroscience 379, 93– 102. Heaton, R.K., Chelune, G.J., T alley , J.L., Kay , G.G., Cur- tiss, G., 1993. WCST: Wisconsin card sorting test. Psy- chological Assessment resources. Hinton, G.E., 2002. T raining pro ducts of exp erts by min- imizing contrastiv e div ergence. Neural computation 14, 1771–1800. Hinton, G.E., 2012. A practical guide to training restricted boltzmann machines, in: Neural netw orks: T ric ks of the trade. Springer, pp. 599–619. Hinton, G.E., Osindero, S., T eh, Y.W., 2006. A fast learning algorithm for deep b elief nets. Neural computation 18, 1527–1554. Hochreiter, S., Sc hmidhuber, J., 1997. Long short-term memory . Neural computation 9, 1735–1780. Hohwy , J., Seth, A., 2020. Predictive pro cessing as a sys- tematic basis for identifying the neural correlates of con- sciousness. Philosophy and the Mind Sciences 1. Holcman, D., Tso dyks, M., 2006. The emergence of up and down states in cortical netw orks. PLoS Computational Biology 2, e23. doi: 10.1371/journal.pcbi.0020023 . Holland, O., 2007. A strongly embo died approach to machine consciousness. Journal of Consciousness Studies 14, 97– 110. Houk, J.C., Davids, J.L., Beiser, D.G. (Eds.), 1995. Mo dels of Information Pro cessing in the Basal Ganglia. The MIT Press, Cambridge, MA. 35 Huang, M.H., Rust, R., Maksimovic, V., 2019. The feeling economy: Managing in the next generation of artificial intelligence (ai). California Management Review 61, 43– 65. Huddy , V., Mansell, W., 2023. Loss and restoration of con- trol: A p erceptual control theory p erspective on the role of mental sim ulation, in: The Interdisciplinary Handbo ok of Perceptual Control Theory , V olume I I. Elsevier, pp. 59–74. Hutto, D.D., 2005. Knowing what? radical versus conserv a- tive enactivism. Phenomenology and the Cognitive Sci- ences 4, 389–405. Jacquey , L., Baldassarre, G., Santucci, V.G., O’Regan, J.K., 2019. Sensorimotor contingencies as a key drive of de- velopmen t: from babies to robots . F rontiers in Neuro- robotics 13, e1–20. doi: 10.3389/fnbot.2019.00098 . Jaeger, H., 2001. The ‘echo state’ approac h to analysing and training recurren t neural net works-with an erratum note. GMD Rep ort 48. German National Research Center for Information T echnology . Bonn, Germany . J¨ an tsch, M., Wittmeier, S., Knoll, A., 2010. Distributed con- trol for an anthropomimetic rob ot, in: 2010 IEEE/RSJ International Conference on Intelligen t Rob ots and Sys- tems, IEEE. pp. 5466–5471. Jeannerod, M., 2003. The mechanism of self-recognition in humans. Behavioural brain researc h 142, 1–15. Jung, M., Matsumoto, T., T ani, J., 2019a. Goal-directed behavior under v ariational predictive coding: Dynamic organization of visual attention and w orking memory , in: 2019 IEEE/RSJ International Conference on Intelligen t Robots and Systems (IROS), IEEE. pp. 1040–1047. Jung, M., Matsumoto, T., T ani, J., 2019b. Goal-Directed Behavior under V ariational Predictive Co ding: Dynamic Organization of Visual Atten tion and W orking Memory . Preprint arXiv 1903.04932v1. Kabat-Zinn, J., 1990. F ull catastrophe living: Using the wisdom of your b ody and mind to face stress, pain, and illness. Delta T rade Paperbacks. New edition 2013, with prev ace b y Thich Nhat Hanh. Kappel, D., Nessler, B., Maass, W., 2014. Stdp installs in winner-take-all circuits an online appro ximation to hidden marko v mo del learning. PLoS Comput Biol 10, e1003511. doi: 10.1371/journal.pcbi.1003511 . Kingma, D.P ., W elling, M., 2013. Auto-encoding v ariational bay es. arXiv . Koch, C., Massimini, M., Boly , M., T ononi, G., 2016. Neural correlates of consciousness: progress and problems. Na- ture Reviews Neuroscience 17, 307. Koch, C., Tsuchiy a, N., 2007. A ttention and consciousness: tw o distinct brain processes. T rends in cognitive sciences 11, 16–22. Kohonen, T., 2001. Self-organizing maps. Third ed., Springer, Berlin. Konidaris, G., Kaelbling, L.P ., Lozano-Perez, T., 2018. F rom skills to sym b ols: Learning symbolic representations for abstract high-level planning. Journal of Artificial Intelli- gence Research 61, 215–289. Kugele, S., F ranklin, S., 2021. Learning in lida. Cognitive Systems Research 66, 176–200. Kumaran, D., Maguire, E.A., 2007. Matc h mismatc h pro- cesses underlie h uman hipp ocampal resp onses to associa- tive novelt y . J Neurosci 27, 8517–8524. Lake, B.M., Ullman, T.D., T enenbaum, J.B., Gershman, S.J., 2017. Building machines that learn and think lik e people. Brain and Beha vioural Sciences 40, 1–72. Lau, H., Rosenthal, D., 2011. Empirical support for higher- order theories of conscious aw areness. T rends in cognitiv e sciences 15, 365–373. LeDoux, J.E., Bro wn, R., 2017. A higher-order theory of emotional consciousness. Pro ceedings of the National Academy of Sciences 114, E2016–E2025. Levy , R., 2012. Apath y: a pathology of goal-directed be- haviour. a new concept of the clinic and pathophysiology of apathy . Revue neurologique 168, 585–597. Levy , R., 2021. A pathology of goal-directed b eha viour and prefrontal cortex–basal ganglia circuits. Apathy: Clini- cal and Neuroscientific Perspectives from Neurology and Psychiatry , 156. Lewis, B.L., 2012. In the game: The interface betw een wat- son and jeopardy! IBM Journal of Research and Devel- opment 56, 17–1. Li, J., Li, D., Xiong, C., Hoi, S., 2022. Blip: Bootstrapping language-image pre-training for unified vision-language understanding and generation, in: In ternational Confer- ence on Machine Learning, PMLR. pp. 12888–12900. Lisman, J.E., Grace, A.A., 2005. The hippocampal-vta lo op: controlling the en try of information into long-term mem- ory . Neuron 46, 703–713. Luko ˇ seviˇ cius, M., Jaeger, H., 2009. Reservoir computing ap- proaches to recurrent neural net work training. Computer science review 3, 127–149. Lynn, C.W., Bassett, D.S., 2019. The physics of brain net- work structure, function and control. Nature Reviews Physics 1, 318–332. Maass, W., Natschl¨ ager, T., Markram, H., 2002. Real-time computing without stable states: a new framework for neural computation based on perturbations. Neural Com- put 14, 2531–2560. doi: 10.1162/089976602760407955 . Malinowski, P ., 2013. Neural mec hanisms of atten tional con- trol in mindfulness meditation. F rontiers in neuroscience 7, 8. Mannella, F., Mirolli, M., Baldassarre, G., 2016. Goal- directed behavior and instrumen tal dev aluation: A neural system-level computational model. F rontiers in Beha v- ioral Neuroscience 10, e1–27. doi: 10.3389/fnbeh.2016. 00181 . Mannella, F., Santucci, V.G., Eszter, S., Jacquey , L., O’Regan, K.J., Baldassarre, G., 2018. Know y our b ody through in trinsic goals. F rontiers in neurorobotics 12, E1– 17. doi: 10.3389/fnbot.2018.00030 . Marcus, G., Davis, E., 2019. Reb ooting AI: Building ar- tificial in telligence we can trust. Pan theon Bo oks, New Y ork. Marques, H.G., Holland, O., 2009. Arc hitectures for func- tional imagination. Neuro computing 72, 743–759. Mars, R.B., Sallet, J., Rushw orth, M.F., Y eung, N. (Eds.), 2011. Neural basis of motiv ational and cognitive control. MIT Press. Marsella, S., Gratch, J., Petta, P ., 2010. Computational models of emotion, in: A Blueprin t for Affective Comput- ing - A sourcebo ok and man ual. Oxford Universit y Press, New Y ork, NY, USA. volume 11, pp. 21–46. Mashour, G.A., Roelfsema, P ., Changeux, J.P ., Dehaene, S., 2020. Conscious pro cessing and the global neuronal workspace hypothesis. Neuron 105, 776–798. doi: 10.1016/ j.neuron.2020.01.026 . Matsumoto, T., Ohata, W., Benureau, F.C., T ani, J., 2022. Goal-directed planning and goal understanding by ex- tended active inference: Ev aluation through simulated and physical rob ot exp erimen ts. Entrop y 24, 469. 36 Mattera, A., P agani, M., Baldassarre, G., 2020. A computa- tional model in tegrating multiple phenomena on cued fear conditioning, extinction, and reinstatement. F rontiers in systems neuroscience 14, 65. McClelland, James L. andRumelhart, D.E., the PDPRe- searchGroup, 1986. Parallel distributed pro cessing: ex- plorations in the microstructure of cognition. v olume 1. The MIT Press, Cambridge,MA. McMurtray , A., Tseng, B., Diaz, N., Chung, J., Meh ta, B., Saito, E., 2014. Acute psychosis asso ciated with sub cor- tical stroke: comparison b et ween basal ganglia and mid- brain lesions. Case Reports in Neurological Medicine 2014. Melloni, L., Mudrik, L., Pitts, M., Ko ch, C., 2021. Making the hard problem of consciousness easier. Science 372, 911–912. Memmert, D., 2009. Pa y attention! a review of visual atten- tional exp ertise in sp ort. International Review of Sp ort and Exercise Psychology 2, 119–138. Meneguzzo, P ., Tsakiris, M., Schioth, H.B., Stein, D.J., Brooks, S.J., 2014. Subliminal versus supraliminal stim- uli activ ate neural resp onses in anterior cingulate cortex, fusiform gyrus and insula: a meta-analysis of fMRI stud- ies. BMC Psychol 2, 52. Metzinger, T., 2013. The m yth of cognitive agency: sub- personal thinking as a cyclically recurring loss of mental autonomy . F rontiers in psyc hology 4, 931. Metzinger, T.K., 2017. The problem of men tal action, in: Metzinger, T.K., Wiese, W. (Eds.), Philosophy and Predictive Pro cessing. MIND Group, F rankfurt am Main. chapter 19. URL: https://predictive- mind.net/ papers/the- problem- of- mental- action , doi: 10.15502/ 9783958573208 . Meyer, K., Damasio, A.R., 2009. Convergence and div er- gence in a neural arc hitecture for recognition and memory . T rends Neurosci 32, 376–382. Miikkulainen, R., Bednar, J.A., Cho e, Y., Sirosh, J., 2006. Computational maps in the visual cortex. Springer Sci- ence & Business Media. Miles, S., Ho wlett, C.A., Berryman, C., Nedeljko vic, M., Moseley , G.L., Phillipou, A., 2021. Considerations for using the wisconsin card sorting test to assess cognitive flexibility . Beha vior research methods 53, 2083–2091. Miller, B.T., Clapp, W.C., 2011. F rom vision to decision: the role of visual atten tion in elite sports p erformance. Eye & contact lens 37, 131–139. Mirolli, M., Mannella, F., Baldassarre, G., 2010. The roles of the amygdala in the affectiv e regulation of bo dy , brain, and behaviour. Connection Science 22, 215–245. Mysore, S.P ., Kothari, N.B., 2020. Mechanisms of competi- tive selection: A canonical neural circuit framew ork. Elife 9. Nair, A., P ong, V., Dalal, M., Bahl, S., Lin, S., Levine, S., 2018. Visual reinforcemen t learning with imag- ined goals, in: The Second Lifelong Learning: A Rein- forcement Learning Approac h W orkshop (LLRLA2018 at F AIM2018). 14/07/2018, Stockholm, Sw eden. Newman, S.D., Carp enter, P .A., V arma, S., Just, M.A., 2003. F rontal and parietal participation in problem solv- ing in the tow er of london: fmri and computational mo d- eling of planning and high-level p erception. Neuropsy- chologia 41, 1668–1682. Oddi, A., Rasconi, R., Santucci, V.G., Sartor, G., Cartoni, E., Mannella, F., Baldassarre, G., 2019. An intrinsi- cally motiv ated planning architecture for curiosity-driv en robots., in: AIR O@ AI*IA, pp. 19–24. Ohlsson, S., 1992. Information-pro cessing explanations of insight and related phenomena, in: Keane, M., Gilho oly , K. (Eds.), Adv ances in the Psychology of Thinking. Harvester-Wheatsheaf. O’Regan, J.K., Noe, A., 2001. A sensorimotor account of vision and visual consciousness. Behav Brain Sci 24, 939– 73; discussion 973–1031. O’Reilly , J.X., Sch¨ uffelgen, U., Cuell, S.F., Behrens, T.E., Mars, R.B., Rushw orth, M.F., 2013. Disso ciable effe cts of surprise and model update in parietal and anterior cin- gulate cortex. Proceedings of the National Academy of Sciences , 201305373. OReilly , R.C., F rank, M.J., 2006. Making working memory work: a computational mo del of learning in the prefrontal cortex and basal ganglia. Neural computation 18, 283– 328. Oudeyer, P .Y., Kaplan, F., Hafner, V.V., 2007. Intrinsic motiv ation systems for autonomous mental development. IEEE transactions on evolutionary computation 11, 265– 286. doi: 10.1109/tevc.2006.890271 . O’Regan, J.K., Myin, E., No¨ e, A., 2005. Sensory conscious- ness explained (better) in terms of corp orality and alerting capacity . Phenomenology and the Cognitive Sciences 4, 369–387. Paiv a, A., Leite, I., Rib eiro, T., 2012. Emotion mo delling for social rob ots, Oxford: Oxford Universit y Press. Panksepp, J., 1998. Affective neuroscience: the foundations of human and animal emotions. Oxford Unv ersit y Press, Oxford. Park, J.S., O’Brien, J.C., Cai, C.J., Morris, M.R., Liang, P ., Bernstein, M.S., 2023. Generative Agents: Interac- tive Sim ulacra of Human Beha vior. T ec hnical Report 2304.03442. arXiv. doi: 10.48550/ARXIV.2304.03442 . Parks, E.L., Madden, D.J., 2013. Brain connectivity and visual attention. Brain Connectivity 3, 317–338. Pasquali, A., Timmermans, B., Cleeremans, A., 2010. Know thyself: Metacognitiv e netw orks and measures of con- sciousness. Cognition 117, 182–190. Passingham, R.E., Wise, S.P ., 2012. The neurobiology of the prefrontal cortex: anatomy , evolution, and the origin of insight. 50, Oxford Universit y Press. Patai, E.Z., Spiers, H.J., 2021. The Versatile Wayfinder: Prefrontal Contributions to Spatial Navigation. T rends Cogn Sci 25, 520–533. doi: 10.1016/j.tics.2021.02.010 . Paus, T., 2001. Primate anterior cingulate cortex: where motor control, drive and cognition interface. Nat Rev Neurosci 2, 417–424. Pennartz, C.M., 2015. The brain’s representational p ow er: on consciousness and the integration of mo dalities. MIT Press. Pennartz, C.M., 2018. Consciousness, representation, action: the importance of being goal-directed. T rends in cognitiv e sciences 22, 137–153. Pennartz, C.M., 2022. What is neurorepresen tationalism? from neural activity and predictive pro cessing to m ulti- level representations and consciousness. Behavioural Brain Research 432, 113969. Pessoa, L., 2015. Multiple influences of reward on perception and attention. Visual cognition 23, 272–290. Pezzulo, G., 2014. Wh y do y ou fear the b ogeyman? an embodied predictiv e coding mo del of perceptual inference. Cognitive, Affectiv e, & Behavioral Neuroscience 14, 902– 911. Pezzulo, G., Rigoli, F., F riston, K., 2015. Active inference, homeostatic regulation and adaptive behavioural control. 37 Progress in neurobiology 134, 17–35. Pfeiffer, B.E., F oster, D.J., 2013. Hipp ocampal place-cell se- quences depict future paths to remem b ered goals. Nature 497, 74. Pitts, M.A., Lutsysh yna, L.A., Hillyard, S.A., 2018. The relationship betw een atten tion and conscious- ness: an expanded taxonomy and implications for ‘no- report’paradigms. Philosophical T ransactions of the Ro yal Society B: Biological Sciences 373, 20170348. P¨ oppel, E., Held, R., F rost, D., 1973. Residual visual func- tion after brain w ounds in volving the cen tral visual path- wa ys in man. Nature 243, 295–296. Pourpanah, F., Abdar, M., Luo, Y., Zhou, X., W ang, R., Lim, C.P ., W ang, X.Z., W u, Q.M.J., 2022. A review of generalized zero-shot learning metho ds , 1–20URL: https://doi.org/10.1109/TPAMI.2022.3191696 , doi: 10. 1109/tpami.2022.3191696 . Po w ers, W.T., 2016. Perceptual Control Theory: An Overview of the Third Grand Theory in Psycho l- ogy—Introductions, Readings, and Resources. Living Control Systems Publ. Prescott, T.J., Gonzalez, F.M.M., Gurney , K., Humphries, M.D., Redgra ve, P ., 2006. A robot model of the basal gan- glia: b ehavior and intrinsic pro cessing. Neural Netw orks 19, 31–61. doi: 10.1016/j.neunet.2005.06.049 . Price, A., Filoteo, J.V., Maddox, W.T., 2009. Rule-based category learning in patients with parkinson’s disease. Neuropsychologia 47, 1213–1226. Raffone, A., Srinivasan, N., v an Leeuw en, C., 2014a. The in- terplay of attention and consciousness in visual searc h, at- tentional blink and w orking memory consolidation. Philo- sophical T ransactions of the Ro yal Society B: Biological Sciences 369, 20130215. Raffone, A., Sriniv asan, N., v an Leeuw en, C., 2014b. P ercep- tual aw areness and its neural basis: bridging exp erimen tal and theoretical paradigms. Philos T rans R So c Lond B Biol Sci 369, 20130203. Raffone, A., Sriniv asan, N., v an Leeuw en, C., 2015. Rapid switching and complementary evidence accumulation en- able flexibilit y of an all-or-none global w orkspace for con- trol of attentional and conscious pro cessing: a reply to wyble et al. Philos T rans R So c Lond B Biol Sci 370, 20140315. Rao, R.P ., Ballard, D.H., 1999. Predictive co ding in the visual cortex: a functional interpretation of some extra- classical rec eptiv e-field effects. Nature Neuroscience 2, 79– 87. doi: 10.1038/4580 . Redgrav e, P ., Prescott, T.J., Gurney , K., 1999. The basal ganglia: a v ertebrate solution to the selection problem? Neuroscience 89, 1009–1023. Reggia, J.A., 2013. The rise of mac hine consciousness: Studying consciousness with computational mo dels. Neu- ral Netw orks 44, 112–131. Rehder, E., Wirth, F., Lauer, M., Stiller, C., 2018. Pedes- trian prediction by planning using deep neural netw orks, in: 2018 IEEE International Conference on Robotics and Automation (ICRA), IEEE. pp. 1–5. Revonsuo, A., 2006. Inner presence: Consciousness as a biological phenomenon. Mit Press. Ribas-F ernandes, J.J.F., Solw ay , A., Diuk, C., McGuire, J.T., Barto, A.G., Niv, Y., Botvinick, M.M., 2011. A neu- ral signature of hierarchical reinforcemen t learning. Neu- ron 71, 370–379. doi: 10.1016/j.neuron.2011.05.042 . Rizzolatti, G., Matelli, M., 2003. Two different streams form the dorsal visual system: anatom y and functions. Exp Brain Res 153, 146–157. Rohaut, B., Doyle, K.W., Reynolds, A.S., Igwe, K., Couch, C., Matory , A., Rizvi, B., Roh, D., V elazquez, A., Megjhani, M., et al., 2019. Deep structural brain lesions associated with consciousness impairment early after hem- orrhagic stroke. Scientific reports 9, 1–9. Rolls, E.T., 2004. The functions of the orbitofron tal cor- tex. Brain Cogn 55, 11–29. doi: 10.1016/S0278- 2626(03) 00277- X . Rueck ert, E., Kappel, D., T anneberg, D., Pecevski, D., Pe- ters, J., 2016. Recurrent spiking netw orks solve plan- ning tasks. Scientific Reports 6, 21142. doi: 10.1038/ srep21142 . Russell, S.J., Norvig, P ., 2016. Artificial Intelligence: A Modern Approac h. Third edition ed., Pearson Education, Harlow, UK. Ryan, R.M., Deci, E.L., 2000. Self-determination theory and the facilitation of in trinsic motiv ation, social development, and well-being. Am Psychol 55, 68–78. Santucci, V.G., Baldassarre, G., Mirolli, M., 2016. Grail: A goal-disco vering rob otic architecture for intrinsically- motiv ated learning. IEEE T ransactions on Cognitiv e and Developmen tal Systems 8, 214–231. Scherer, K.R., 2005. What are emotions? and how can they be measured? So cial science information 44, 695–729. doi: 10.1177/0539018405058216 . Schmidh uber, J., 1991. A p ossibilit y for implemen ting cu- riosity and boredom in model-building neural con trollers, in: Pro c. of the international conference on sim ulation of adaptive behavior: F rom animals to animats, pp. 222–227. Schooler, J.W., Smallwood, J., Christoff, K., Handy , T.C., Reichle, E.D., Say ette, M.A., 2011. Meta-aw areness, p er- ceptual decoupling and the wandering mind. T rends in cognitive sciences 15, 319–326. Schroll, H., Hamk er, F.H., 2013. Computational models of basal-ganglia pathw ay functions: focus on functional neuroanatomy . F rontiers in System Neuroscience 7, 122. doi: 10.3389/fnsys.2013.00122 . Sch ultz, W., 2002. Getting formal with dopamine and re- ward. Neuron 36, 241–263. Seger, C.A., 2008. How do the basal ganglia contribute to categorization? their roles in generalization, resp onse se- lection, and learning via feedback. Neuroscience & Biobe- havioral Reviews 32, 265–278. Seth, A.K., Bayne, T., 2022. Theories of consciousness. Na- ture Reviews Neuroscience , 1–14. Seth, A.K., Izhikevic h, E., Reeke, G.N., Edelman, G.M., 2006. Theories and measures of consciousness: an ex- tended framew ork. Proc Natl Acad Sci U S A 103, 10799– 10804. Simon, H.A., 1975. The functional equiv alence of problem solving skills. Cognitive psyc hology 7, 268–288. Soltani, A., Koechlin, E., 2022. Computational models of adaptive behavior and prefrontal cortex. Neuropsy- chopharmacology 47, 58–71. Sriv asta v a, A., al., 2022. Beyond the Imitation Game: Quantifying and extrapolating the capabilities of language models. T echnical Report. . Stumbrys, T., Erlacher, D., Sc h¨ adlic h, M., Sc hredl, M., 2012. Induction of lucid dreams: A systematic review of evi- dence. Consciousness and Cognition 21, 1456–1475. Sun, R., 2016. The clarion cognitive architecture: T o ward a comprehensiv e theory of the mind, Oxford Univ ersity Press. chapter 6, pp. 117–133. Sun, R., Wilson, N., Lynch, M., 2016. Emotion: A unified 38 mechanistic interpretation from a cognitive architecture. Cognitive Computation 8, 1–14. Sutton, R.S., Barto, A.G., 1998. In tro duction to reinforce- ment learning. volume 135. MIT press Cambridge. T ang, W., Shin, J.D., Jadhav, S.P ., 2021. Multiple time- scales of decision-making in the hipp ocampus and pre- frontal cortex. Elife 10, e66227. T ang, Y.Y., Holzel, B.K., Posner, M.I., 2015. The neuro- science of mindfulness meditation. Nature Reviews Neu- roscience 16, 213–225. doi: 10.1038/nrn3916 . T egmark, M., 2017. Life 3.0: Being human in the age of artificial intelligence. Knopf. T elles-Correia, D., Moreira, A.L., Goncalv es, J.S., 2015. Hal- lucinations and related concepts-their conceptual back- ground. F rontiers in psyc hology 6, 991. Thill, S., Caligiore, D., Borghi, A.M., Ziemk e, T., Baldas- sarre, G., 2013. Theories and computational mo dels of af- fordance and mirror systems: An integrativ e review. Neu- roscience and Biobehavioral Reviews 37, 491–521. T ononi, G., 2004. An information in tegration theory of con- sciousness. BMC Neurosci 5, 42. T ononi, G., 2008. Consciousness as integrated information: a provisional manifesto. Biol Bull 215, 216–242. T ononi, G., Boly , M., Massimini, M., Koch, C., 2016. Inte- grated information theory: from consciousness to its phys- ical substrate. Nat Rev Neurosci 17, 450–461. Tsujimoto, S., Genovesio, A., Wise, S.P ., 2011. F rontal pole cortex: encoding ends at the end of the endbrain. T rends in cognitive sciences 15, 169–176. Ty e, K.M., 2018. Neural Circuit Motifs in V alence Process- ing. Neuron 100, 436–452. doi: 10.1016/j.neuron.2018. 10.001 . Ullman, S., 2019. Using neuroscience to dev elop artificial intelligence. Science 363, 692–693. V an Boxtel, J.J., Tsuchiy a, N., Ko c h, C., 2010. Conscious- ness and attention: on sufficiency and necessit y . F rontiers in Psychology 1, 217. V aswani, A., Shazeer, N., Parmar, N., Uszkoreit, J., Jones, L., Gomez, A.N., Kaiser, L., Polosukhin, I., 2017. At- tention is all you need. arXiv preprint . V erdejo-Garc ´ ıa, A., P´ erez-Garc ´ ıa, M., Bechara, A., 2006. Emotion, decision-making and substance dep endence: a somatic-marker model of addiction. Curr Neuropharmacol 4, 17–31. V oss, U., Holzmann, R., T uin, I., Hobson, A.J., 2009. Lucid dreaming: a state of consciousness with features of both waking and non-lucid dreaming. Sleep 32, 1191–1200. V ossel, S., Geng, J.J., Fink, G.R., 2014. Dorsal and ven tral attention systems: distinct neural circuits but collab ora- tive roles. The Neuroscien tist 20, 150–159. W ang, Y., Zuo, C., W ang, D., T ao, S., Hao, L., 2020. Reduced thalamus volume and enhanced thalamus and fronto-parietal netw ork integration in the chess exp erts. Cerebral Cortex 30, 5560–5569. W ard, P ., Ca v anna, A.E., et al., 2013. F unctional neu- roanatomy and b eha vioural correlates of the basal gan- glia: evidence from lesion studies. Behavioural neurology 26, 219–223. W ayne, G., Hung, C.C., Amos, D., Mirza, M., Ah uja, A., Grabsk a-Barwinsk a, A., Rae, J., Miro wski, P ., Leib o, J.Z., Santoro, A., Gemici, M., Reynolds, M., Harley , T., Abramson, J., Mohamed, S., Rezende, D., Saxton, D., Cain, A., Hillier, C., Silver, D., Ka vukcuoglu, K., Botvinick, M., Hassabis, D., Lillicrap, T., 2018. Unsup er- vised predictive memory in a goal-directed agent. arXiv preprint arXiv:1803.10760 . W eiskrantz, L., 1991. Disconnected aw areness for detect- ing, processing, and remembering in neurological patien ts. Journal of the Roy al Society of Medicine 84, 466–470. W eiskrantz, L., 1995. The problem of animal consciousness in relation to neuropsyc hology . Beha vioural brain research 71, 171–175. W eiskrantz, L., 2004. Ro ots of blindsight. Progress in brain research 144, 227–241. W o darz, N., Beck er, T., Dec kert, J., 1995. Musical halluci- nations associated with post-thyroidectom y h yp oparath y- roidism and symmetric basal ganglia calcifications. Jour- nal of Neurology , Neurosurgery , and Psychiatry 58, 763. Y aron, I., Melloni, L., Pitts, M., Mudrik, L., 2022. The contrast database for analysing and comparing empiri- cal studies of consciousness theories. Nature Human Be- haviour 6, 593–604. Y ates, J.C., Immergut, M., 2015. The Mind Illuminated: A Complete Meditation Guide In tegrating Buddhist Wis- dom and Brain Science. Y un, T., Sun, C., Pa vlick, E., 2021. Does Vision- and-Language Pretraining Impro ve Lexical Grounding? preprint 2109.10246. arXiv. v an der Zw aard, R., Polak, M.A., 2001. Pseudohallucina- tions: a pseudoconcept? a review of the v alidity of the concept, related to asso ciated symptomatology . Compre- hensive Psychiatry 42, 42–50. 39
Original Paper
Loading high-quality paper...
Comments & Academic Discussion
Loading comments...
Leave a Comment