Nonlinear and Nonlocal Elasticity in Coarse-Grained Differential-Tension Models of Epithelia
The shapes of epithelial tissues result from a complex interplay of contractile forces in the cytoskeleta of the cells in the tissue, and adhesion forces between them. A host of discrete, cell-based models describe these forces by assigning different surface tensions to the apical, basal, and lateral sides of the cells. These differential-tension models have been used to describe the deformations of epithelia in different living systems, but the underlying continuum mechanics at the scale of the epithelium are still unclear. Here, we derive a continuum theory for a simple differential-tension model of a two-dimensional epithelium and study the buckling of this epithelium under imposed compression. The analysis reveals how the cell-level properties encoded in the differential-tension model lead to linear, nonlinear as well as nonlocal elastic behavior at the continuum level.
💡 Research Summary
The paper presents a systematic derivation of a continuum theory from a simple differential‑tension model of a two‑dimensional epithelial sheet composed of isosceles trapezoidal cells. Each cell has three distinct surface tensions: apical (Γₐ), basal (Γ_b) and lateral (Γ). The discrete energy of a single cell is E = ΓₐLₐ + Γ_bL_b + ΓL, where Lₐ, L_b and L are the lengths of the corresponding sides. By non‑dimensionalising with the square root of the cell area (√A_c) the authors introduce dimensionless parameters λ = L/√A_c, κ = K/√A_c (mean base), and the ratios α = Γₐ/Γ, β = Γ_b/Γ`. The differential tension is captured by δ = α – β, while the dimensionless thickness of the flat epithelium is ℓ₀ = 1/λ₀.
In the continuum limit the angle φ(s) describing the tilt of each cell becomes a smooth function of the arclength s of the undeformed midline. The total energy is written as an integral E = ∫ e(φ) ds with the energy density e(φ) = ℓ₀² λ secφ + λ(1 + δ sinφ). The authors assume a scaling regime where λ = O(ℓ₀) and φ = O(ℓ₀⁻²), which allows cells to undergo small but finite deformations while preserving area. Under these scalings the relation between φ and the midline tangent angle ψ(s) is expressed through an infinite series involving Bernoulli numbers. By inverting this series order‑by‑order they obtain φ as a power series in ψ and its derivatives: φ = ψ’/(2ℓ₀) – ψ’’’/(24ℓ₀³) + … . This expansion goes beyond previous work that kept only the first term, thereby revealing higher‑order nonlinear and nonlocal contributions.
Projecting the discrete geometry onto the Cartesian axes yields differential relations for the centreline coordinates (x(s), y(s)). After expanding in powers of ℓ₀⁻¹ the authors find dx/ds = f cosψ – g sinψ, dy/ds = f sinψ + g cosψ, with f = 1 + ψ’²/(24ℓ₀²) + … and g = ψ’’/(12ℓ₀²) + … . The term g, absent in the classical Euler‑Elastica, encodes nonlocal elasticity because it involves higher spatial derivatives of ψ.
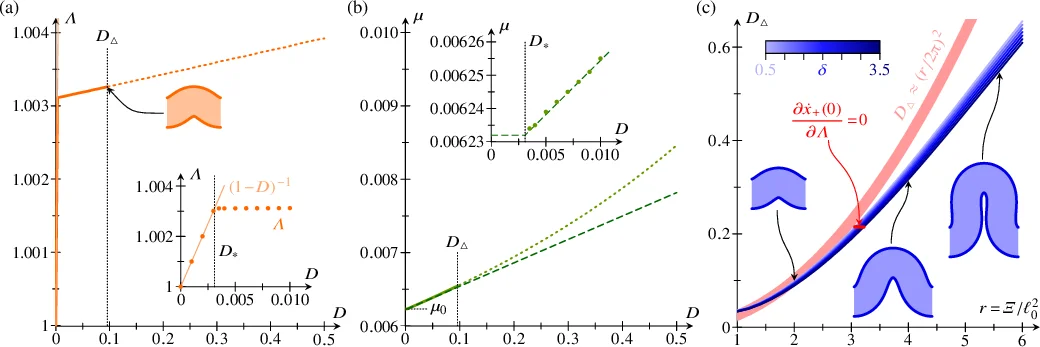
To study buckling under imposed compression, a horizontal compressive force μ is introduced as a Lagrange multiplier enforcing a prescribed end‑to‑end shortening D. The Lagrangian is expanded in the small parameter Ξ = ℓ₀ Σ (Σ is half the undeformed length), which is proportional to the number of cells N = 2Ξ. Truncating the expansion at fourth order yields a Landau‑type energy containing both nonlinear elastic terms (ψ⁴, ψ’⁴) and nonlocal terms (ψ’‘ψ’’’, ψ’²ψ’’). Variation of this truncated energy leads to a fourth‑order nonlinear differential equation for ψ (Eq. 21). The equation depends on the combined parameter Δ = δℓ₀² and on the dimensionless thickness Λ = λ/ℓ₀, but the nonlocal contributions arise solely from the geometric coupling of cells.
A weakly nonlinear analysis is performed by expanding ψ = ε ψ₀ + ε² ψ₁ + ε³ ψ₂ … with ε² = 1 – Λ(1 – D). At leading order the eigenvalue problem yields μ₀ = z₁ – z, where z = ξ²/4·(1 + ξ²/6) and ξ = πΞ. The corresponding mode is ψ₀(σ) = Ψ₀ sin(πσ). Higher‑order calculations give μ₁ = 0 (indicating a supercritical pitchfork bifurcation) and explicit expressions for ψ₁, ψ₂, and the amplitudes Ψ₀, Ψ₁, Ψ₂ in terms of ξ and Δ. Importantly, the buckling threshold D* = 1 – √(1 – z) emerges independent of the differential tension δ, showing that the onset of instability is governed solely by the geometric stiffness of the sheet.
The post‑buckling energy is then evaluated, revealing that the excess compression d = D – D* leads to an amplitude scaling ε ∝ √d. The final energy expression contains contributions proportional to ε⁴ and to ε²(1 – z)⁻¹, confirming the supercritical nature of the bifurcation. The optimal Λ (i.e., the effective thickness) is obtained by minimizing the total energy with respect to Λ, which couples the geometric compression to the cell‑level tension asymmetry Δ.
Overall, the paper demonstrates that a simple differential‑tension model, when coarse‑grained, yields a continuum description featuring both nonlinear elasticity and nonlocal elasticity arising from the discrete geometry of the cells. The analysis clarifies that while the critical compression for buckling does not depend on the differential tension, the post‑buckling shape, amplitude, and effective stiffness are strongly modulated by δ. This work bridges the gap between cell‑scale mechanical models and tissue‑scale continuum mechanics, providing a rigorous framework for interpreting epithelial folding, morphogenetic buckling, and related phenomena in developmental biology.
Comments & Academic Discussion
Loading comments...
Leave a Comment