Fundamental limits to collective concentration sensing in cell populations
The precision of concentration sensing is improved when cells communicate. Here we derive the physical limits to concentration sensing for cells that communicate over short distances by directly exchanging small molecules (juxtacrine signaling), or over longer distances by secreting and sensing a diffusive messenger molecule (autocrine signaling). In the latter case, we find that the optimal cell spacing can be large, due to a tradeoff between maintaining communication strength and reducing signal cross-correlations. This leads to the surprising result that autocrine signaling allows more precise sensing than juxtacrine signaling for sufficiently large populations. We compare our results to data from a wide variety of communicating cell types.
💡 Research Summary
This paper investigates how communication among cells influences the fundamental limits of chemical concentration sensing. The authors develop a unified physical framework that treats two ubiquitous modes of intercellular signaling: short‑range juxtacrine signaling, where neighboring cells exchange small messenger molecules directly, and long‑range autocrine signaling, where each cell secretes a diffusible messenger that can be sensed by any other cell. Starting from the diffusion equation for an extracellular ligand (diffusivity D_c) and stochastic binding/unbinding dynamics on cell surfaces (rates α and μ), they write linear stochastic differential equations for the ligand field, the bound‑receptor numbers r_i(t), and the messenger populations. Gaussian white noise terms capture both extrinsic fluctuations (ligand diffusion) and intrinsic fluctuations (finite numbers of receptors and messenger molecules).
For a single isolated cell the well‑known Berg‑Purcell limit emerges: the relative error (variance/mean²) in the messenger readout scales as 1/(π a c̄ D_c T) plus terms that vanish when receptor and messenger numbers are large. The analysis then proceeds to coupled cells. In the juxtacrine case the messenger dynamics include production (β r_i), degradation (ν), and exchange with each neighbor at rate γ. A “communication length” λ = 2a γ/ν naturally appears, governing how far a messenger can travel before being degraded. Solving the linear system in Fourier space yields an exact expression for the error for any number of cells N and any geometry. For two cells, the extrinsic part of the error reduces to (3/8)·1/(π a c̄ D_c T) when λ≫a, i.e., strong communication. This is only a modest improvement over the single‑cell case (1/2) because adjacent cells sample overlapping extracellular volumes, creating cross‑correlations that limit independent information gain.
In the autocrine scenario the messenger ρ(x,t) diffuses with coefficient D_ρ and degrades with rate ν, producing a steady‑state concentration profile that decays as e^{-|x−x_i|/λ}/|x−x_i| with λ = D_ρ/ν. Each cell integrates the messenger concentration over its own volume, defining m_i = ∫_{V_i} ρ dV. The error for two cells separated by distance ℓ (ℓ > a) can be written analytically; in the strong‑communication limit (λ≫a) the relative error becomes (2/5)·1/(π a c̄ D_c T ℓ). The error is minimized at an optimal separation ℓ* ≈ 8a/3, where the prefactor 2/5 is the lowest achievable. At this distance the mean messenger signal is reduced (because of the spatial decay) but the variance drops even more sharply, owing to two effects: (i) reduced ligand cross‑correlations between the cells, and (ii) diffusion of the messenger itself smoothing out spatial fluctuations (a “super‑Poissonian” noise reduction).
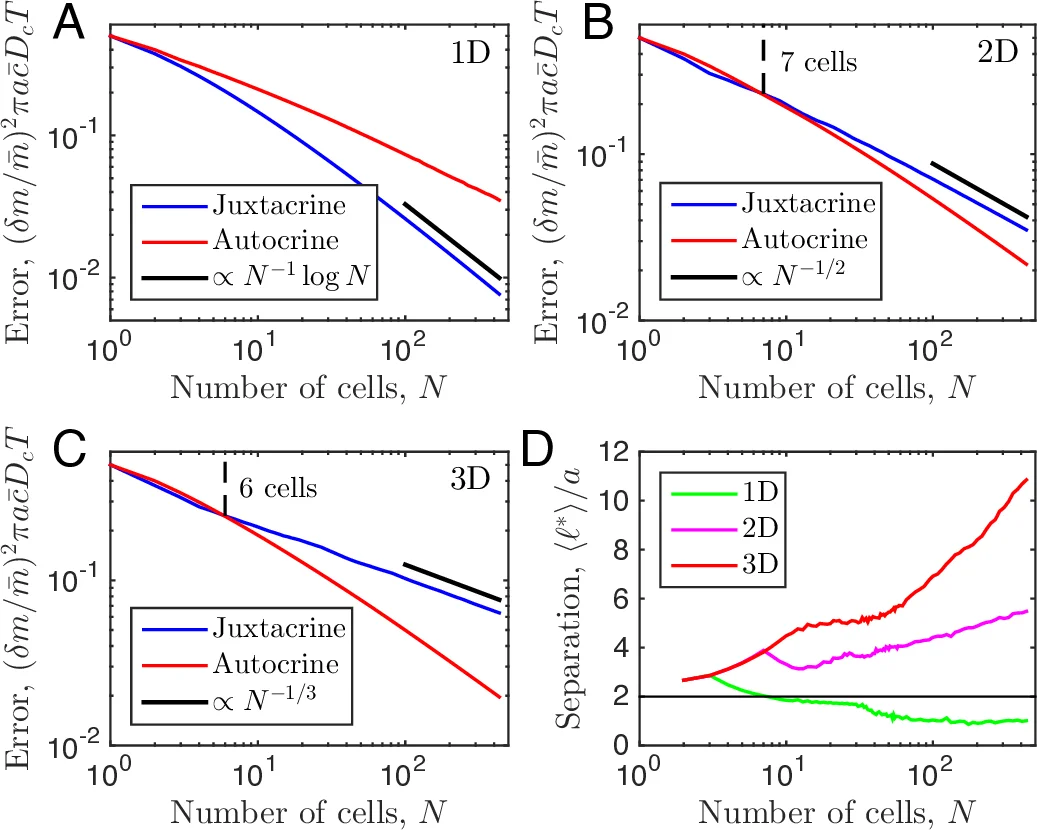
The authors then explore many‑cell configurations in one, two, and three dimensions. For juxtacrine signaling they assume cells occupy a regular lattice with nearest‑neighbor spacing 2a (the densest packing compatible with non‑overlap). In this regime the whole cluster behaves as a single elongated detector; the error scales as N⁻¹ in 1D, N⁻¹/2 in 2D, and N⁻¹/3 in 3D (with a logarithmic correction in 1D), reflecting the inverse dependence on the longest linear dimension of the detector.
For autocrine signaling the optimal arrangement is obtained by Monte‑Carlo minimization of the error while allowing cells to move freely within the given dimensionality. The resulting optimal nearest‑neighbor distance ⟨ℓ*⟩ grows with N, reaching several cell radii for N≈10³–10⁴ in 3D. Consequently, the error decays more steeply than in the juxtacrine case (approximately as N⁻¹·log N in 2D/3D), meaning that for sufficiently large populations autocrine signaling outperforms juxtacrine signaling despite the longer distances between cells.
To connect theory with biology, the authors map experimentally measured parameters onto their phase diagram of communication length λ versus population size N. Bacterial quorum‑sensing systems (λ≈5–20 mm, N≈10²–10⁴) fall in the region where sparse autocrine packing is optimal. Gap‑junction–mediated juxtacrine communication in mammalian tissues (λ≈10–40 µm, N≈10¹–10³) lies in the dense‑packing region where juxtacrine yields the lowest error. They also cite glioblastoma tumor spheroids (N≈10³–10⁴) that secrete IL‑6 and VEGF, which correspond to the autocrine‑optimal sparse regime.
Overall, the paper demonstrates that the intuitive notion “closer cells = better sensing” is incomplete. While short‑range juxtacrine communication benefits from strong coupling, it is limited by shared sampling of the extracellular ligand field. Long‑range autocrine communication can exploit spatial separation to decorrelate noise, and for large 2D/3D populations this yields a fundamentally lower sensing error. These results provide quantitative design principles for natural multicellular systems and for engineered tissues or synthetic cell consortia where precise chemical sensing is required.
Comments & Academic Discussion
Loading comments...
Leave a Comment