Global Optimization, Local Adaptation, and the Role of Growth in Distribution Networks
Highly-optimized complex transport networks serve crucial functions in many man-made and natural systems such as power grids and plant or animal vasculature. Often, the relevant optimization functional is non-convex and characterized by many local extrema. In general, finding the global, or nearly global optimum is difficult. In biological systems, it is believed that natural selection slowly guides the network towards an optimized state. However, general coarse grained models for flow networks with local positive feedback rules for the vessel conductivity typically get trapped in low efficiency, local minima. In this work we show how the growth of the underlying tissue, coupled to the dynamical equations for network development, can drive the system to a dramatically improved optimal state. This general model provides a surprisingly simple explanation for the appearance of highly optimized transport networks in biology such as leaf and animal vasculature.
💡 Research Summary
The paper addresses a fundamental problem in the design of complex transport networks: the underlying optimization functional is typically non‑convex, leading to a rugged landscape with many local minima. While biological distribution systems such as plant leaf venation or animal vasculature are believed to have evolved toward highly efficient configurations, coarse‑grained models that implement only local positive‑feedback rules for vessel conductivity tend to become trapped in suboptimal, poorly organized states.
Ronellenfitsch and Katifori propose a minimal yet powerful extension of these models by explicitly incorporating the growth of the tissue that hosts the network. They consider a two‑dimensional sheet that expands isotropically, with distances scaling as d(t)=λ(t)d(0) and areas as A(t)=λ(t)^2A(0). The flow on an edge e is given by the usual linear relation F_e = K_e (p_j−p_i)/L_e, where K_e is the conductivity and L_e the edge length. Conductivity evolves according to a generalized adaptation rule
dK_e/dt = a (F_e/ĤF)^{2γ} – b K_e + c,
where the first term implements a positive feedback proportional to the squared flow (γ≈2/3 for Poiseuille‑type flow), the second term provides linear decay, and the third term represents a background production of conductivity.
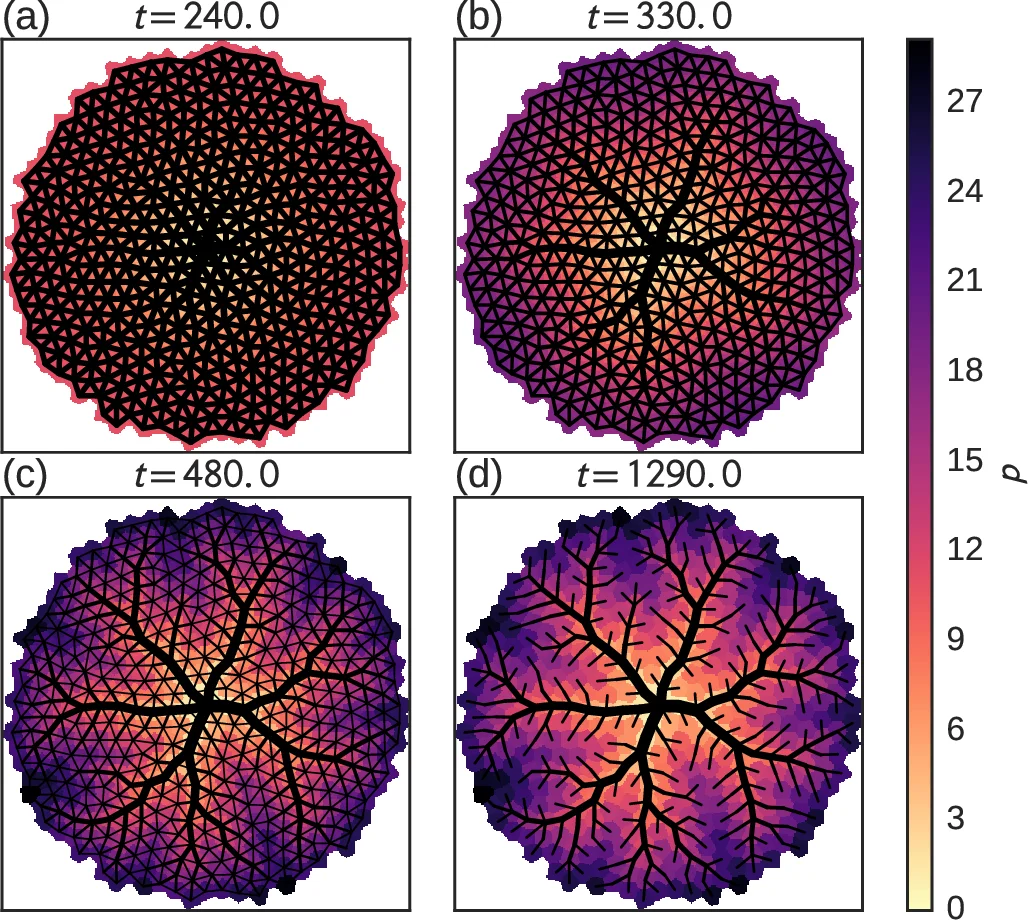
Because the flow itself scales with the growth factor λ(t) (specifically F ∝ λ(t)^δ, with δ≈2 for area‑scaled source terms), the adaptation equation acquires explicit time‑dependent prefactors. In the early exponential growth phase (λ(t)=e^{rt}) the flow term grows exponentially while the background term decays as λ(t)^{-2γδ}. Consequently, the system passes through two distinct dynamical regimes: an initial homogeneous phase dominated by the background term, and a later hierarchical phase where only edges carrying the strongest flows continue to be reinforced.
Two dimensionless parameters control the outcome:
- ρ = b/(rγδ) – the ratio of the intrinsic adaptation timescale to the growth timescale.
- κ = (c/a)(ĤF/ĤS)^{2γ} – the relative strength of background production versus flow‑driven adaptation.
Numerical simulations on disordered tessellations with N≈435 nodes reveal a clear phase diagram in the (ρ,κ) plane. For small ρ·κ the system behaves stochastically: different random initial conductivities lead to a variety of final topologies with a broad distribution of the dissipation functional
E = Σ_e L_e (F_e)^2 / K_e,
and the networks remain largely unstructured. For large ρ·κ the dynamics become deterministic: regardless of the initial condition, the system converges to a unique, highly organized hierarchy. The energy of these deterministic solutions is comparable to the global minima found by computationally intensive simulated‑annealing runs (e.g., E_min≈15.93 versus E_anneal≈15.98 in the authors’ nondimensional units).
Structural analysis shows that the most efficient networks have short mean branch lengths (few edges between bifurcations), reflecting the reuse of major conduits to supply large tissue regions—a hallmark of real leaf venation and arterial trees. The authors also demonstrate that the same optimization effect can be achieved by simply increasing the effective perfusion (λ‑independent) without explicit tissue growth, indicating that the crucial factor is the scaling of flow rather than the mechanical expansion per se.
In summary, the study provides a concise physical mechanism—growth‑induced scaling of flow combined with local positive feedback—that enables a system to self‑organize into near‑globally optimal transport architectures without any global information exchange or stochastic exploration. This insight bridges the gap between evolutionary explanations of vascular optimization and mechanistic models of development, and it suggests new design principles for engineered distribution networks where controlled growth or analogous scaling could be exploited to achieve high efficiency.
Comments & Academic Discussion
Loading comments...
Leave a Comment