Cooperativity and Frustration in Protein-Mediated Parallel Actin Bundles
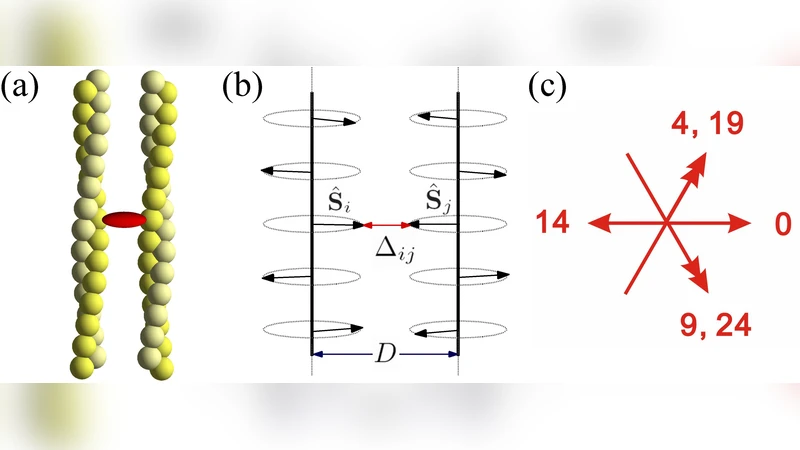
We examine the mechanism of bundling of cytoskeletal actin filaments by two representative bundling proteins, fascin and espin. Small-angle X-ray studies show that increased binding from linkers drives a systematic \textit{overtwist} of actin filaments from their native state, which occurs in a linker-dependent fashion. Fascin bundles actin into a continuous spectrum of intermediate twist states, while espin only allows for untwisted actin filaments and fully-overtwisted bundles. Based on a coarse-grained, statistical model of protein binding, we show that the interplay between binding geometry and the intrinsic \textit{flexibility} of linkers mediates cooperative binding in the bundle. We attribute the respective continuous/discontinous bundling mechanisms of fascin/espin to differences in the stiffness of linker bonds themselves.
💡 Research Summary
This paper investigates how two representative actin‑bundling proteins, fascin and espin, organize parallel actin filaments into higher‑order bundles and how the mechanical properties of the cross‑linking proteins dictate the cooperativity of binding. Using small‑angle X‑ray scattering (SAXS), the authors measured the structural response of actin filaments as a function of increasing concentrations of each bundling protein. The key observation is that binding of either protein induces an “overtwist” of the actin helix away from its native 13/6 symmetry, but the manner of this twist transition differs dramatically between the two proteins. Fascin produces a continuous spectrum of intermediate twist states: as fascin occupancy rises, the SAXS Bragg peak shifts gradually and narrows, indicating a smooth increase in filament pitch and a modest rotation of ~2–3°. In contrast, espin shows a bistable behavior. At low espin concentrations the actin lattice remains essentially untwisted; once a critical espin density is reached, the Bragg peak jumps abruptly to a position corresponding to a highly over‑twisted 28/13 geometry, reflecting a first‑order‑like transition.
To rationalize these findings, the authors develop a coarse‑grained statistical mechanical model that treats actin filaments as a two‑dimensional lattice of binding sites. Each site can be either empty or occupied by a cross‑linker, and the binding energy depends on the local configuration of neighboring sites. Two central parameters capture the physical nature of the linkers: (i) a flexibility coefficient κ that quantifies how much the linker can stretch or bend to accommodate different filament orientations, and (ii) an angular constraint θ that limits the allowable deviation from the native filament geometry. Fascin is modeled with a large κ (high flexibility) and a broad θ (wide angular tolerance), whereas espin is assigned a small κ (rigid) and a narrow θ (tight angular tolerance). Monte‑Carlo simulations of this lattice model reproduce the experimentally observed binding curves. For flexible linkers (high κ), the occupancy increases smoothly with concentration, and the average filament twist follows a continuous function of linker density. For rigid linkers (low κ), the system remains in a low‑twist state until a threshold occupancy is reached, after which a cooperative “all‑or‑none” binding event drives the lattice into the over‑twisted state. The model also predicts that modest changes in κ can convert a continuous transition into a discontinuous one, and vice versa, highlighting the pivotal role of linker stiffness in governing cooperativity.
Beyond the quantitative match, the study offers several mechanistic insights. First, the geometry of the binding interface—how many actin subdomains a linker contacts—determines the degree of geometric frustration that must be resolved during bundling. Flexible linkers can absorb this frustration by allowing slight rotations of the filaments, leading to a spectrum of metastable twist states. Rigid linkers cannot accommodate such strain; instead, they force the system to either remain untwisted or to undergo a collective reorientation that eliminates the frustration in a single step. Second, the cooperative binding mediated by mechanical coupling can be viewed as an effective “allosteric” interaction across the bundle: the state of one linker influences the binding energy of its neighbors through the shared filament lattice. This long‑range coupling is amplified when the linker is stiff, producing the sharp transition observed for espin. Third, the authors discuss the biological relevance of these two modes. Fascin’s continuous response is well suited for dynamic structures such as filopodia, where gradual modulation of bundle stiffness and twist may be required for rapid protrusion and retraction. Espin’s bistable behavior aligns with the need for highly ordered, mechanically robust bundles in sensory hair cells, where a sudden switch to a maximally twisted, tightly packed configuration provides the rigidity needed for mechanotransduction.
In summary, the paper combines high‑resolution SAXS experiments with a minimal yet powerful statistical model to demonstrate that the mechanical stiffness of actin‑binding linkers dictates whether bundling proceeds via a continuous spectrum of twist states (as with fascin) or via a discrete, cooperative transition (as with espin). This work advances our understanding of how protein‑mediated mechanical frustration and cooperativity shape the architecture of the cytoskeleton, and it suggests that tuning linker flexibility could be a general strategy cells employ to regulate bundle mechanics in diverse physiological contexts.
Comments & Academic Discussion
Loading comments...
Leave a Comment