Interplay between microdynamics and macrorheology in vesicle suspensions
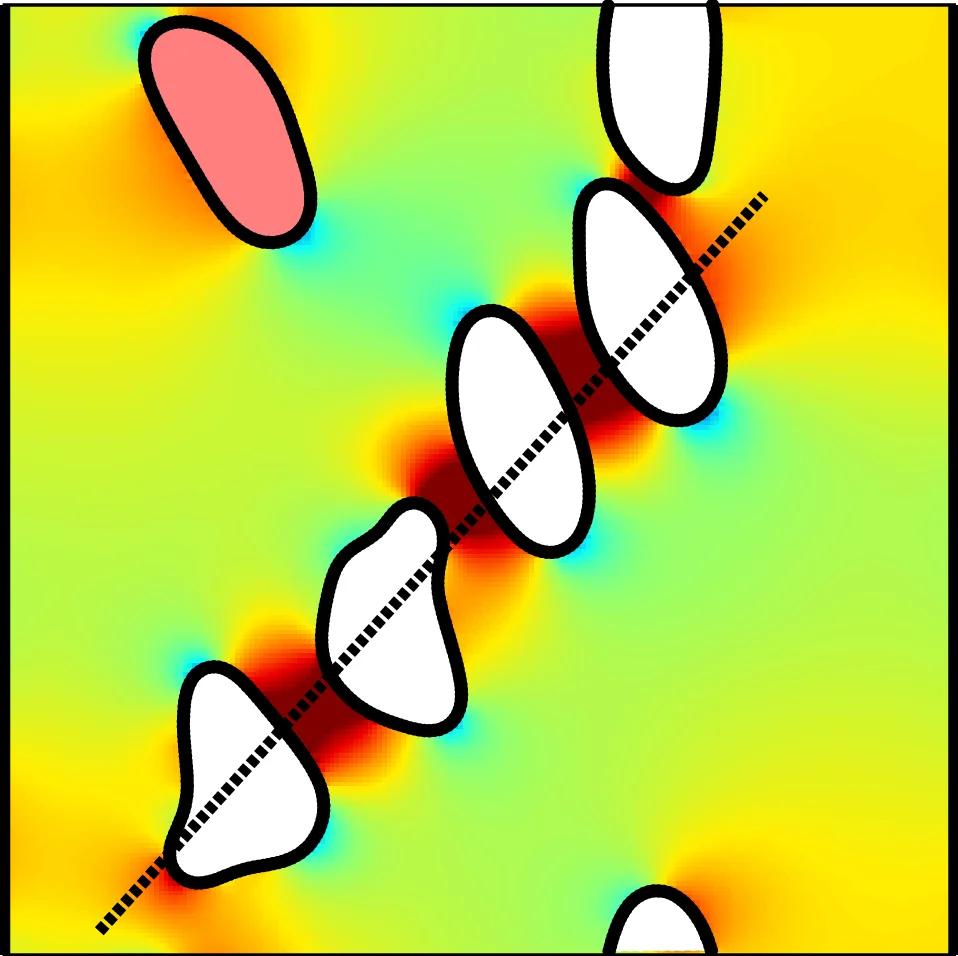
The microscopic dynamics of objects suspended in a fluid determines the macroscopic rheology of a suspension. For example, as shown by Danker and Misbah [Phys. Rev. Lett. {\bf 98}, 088104 (2007)], the viscosity of a dilute suspension of fluid-filled vesicles is a non-monotonic function of the viscosity contrast (the ratio between the viscosities of the internal encapsulated and the external suspending fluids) and exhibits a minimum at the critical point of the tank-treading-to-tumbling transition. By performing numerical simulations, we recover this effect and demonstrate that it persists for a wide range of vesicle parameters such as the concentration, membrane deformability, or swelling degree. We also explain why other numerical and experimental studies lead to contradicting results. Furthermore, our simulations show that this effect even persists in non-dilute and confined suspensions, but that it becomes less pronounced at higher concentrations and for more swollen vesicles. For dense suspensions and for spherical (circular in 2D) vesicles, the intrinsic viscosity tends to depend weakly on the viscosity contrast.
💡 Research Summary
This paper investigates the coupling between microscopic dynamics of vesicles suspended in a fluid and the macroscopic rheology of the resulting suspension. Using a two‑dimensional lattice‑Boltzmann method (LBM) combined with an immersed‑boundary method (IBM), the authors simulate vesicles with varying internal‑to‑external viscosity ratios (viscosity contrast Λ), concentrations φ, membrane rigidity (capillary number Ca), swelling degree Δ, and confinement χ. The study revisits the “Danker‑Misbah effect,” which predicts a non‑monotonic dependence of the intrinsic viscosity η of a dilute vesicle suspension on Λ: η decreases with Λ while vesicles are in the tank‑treading (TT) regime, reaches a minimum at the TT‑to‑tumbling (TB) transition, and then increases in the TB regime.
Key methodological points:
- Fluid dynamics are solved with LBM (D2Q9, single‑relaxation‑time BGK) on a 200 × 200 lattice.
- Vesicle membranes are modeled as inextensible, bending‑elastic curves; forces follow the 2D Helfrich formulation with a Lagrange multiplier enforcing perimeter conservation.
- The IBM interpolates fluid velocities to membrane nodes, advects the membrane, computes elastic forces, and spreads them back to the fluid.
- The effective suspension viscosity η* is obtained from the wall shear stress σ_xy and the imposed shear rate γ; the intrinsic viscosity η = (η* − η₀)/(η₀ φ) isolates the contribution of vesicles.
Results are presented in three stages.
-
Single‑vesicle (dilute limit) case – With Re = 0.5, Ca = 10, Δ = 0.8, and confinement χ = 0.2, the authors examine two representative viscosity contrasts: Λ = 4 (TT) and Λ = 16 (TB). In the TT state the inclination angle θ is steady, the wall stress is constant, and η(t) shows negligible temporal fluctuations. In the TB state θ oscillates periodically, leading to a corresponding oscillation of σ_xy and η(t). Averaging over time yields an intrinsic viscosity η that decreases with Λ in the TT regime, reaches a minimum near Λ_c ≈ 7.8 (the predicted TT‑TB transition for the chosen confinement), and then rises for larger Λ. This reproduces the non‑monotonic trend reported analytically by Danker and Misbah and numerically by Ghigliotti et al., despite the finite Reynolds number and confinement.
-
Effect of vesicle concentration – Simulations are performed for φ = 7.5 %, 15.1 %, and 22.6 % (3, 6, 9 vesicles). As φ increases, vesicle‑vesicle collisions become more frequent. Collisions temporarily increase the inclination angle of the interacting vesicles, producing sharp spikes in the instantaneous intrinsic viscosity. Consequently, η(t) exhibits larger fluctuations and the time‑averaged η grows with φ, reflecting the higher effective solid fraction. Nevertheless, the η(Λ) curve retains its non‑monotonic shape for all concentrations; the minimum becomes less pronounced at the highest φ, indicating that crowding smooths the TT‑TB transition.
-
Influence of membrane rigidity and swelling – Varying the capillary number Ca modifies the vesicle’s deformability. Lower Ca (more flexible membranes) shifts the TT‑TB transition to smaller Λ, moving the viscosity minimum leftward; higher Ca (stiffer membranes) pushes the transition to larger Λ. Changing the swelling degree Δ alters the vesicle’s shape: nearly circular vesicles (Δ → 1) have limited shape change, suppressing the TT‑TB transition and yielding an intrinsic viscosity that is almost independent of Λ. This explains why some experiments and simulations report a monotonic η(Λ) relationship when using highly swollen or nearly spherical particles.
The authors also discuss discrepancies with earlier works that reported monotonic viscosity trends. They argue that differences in numerical implementation (e.g., treatment of collisions, boundary conditions, Reynolds number) can mask the subtle TT‑TB transition, especially in confined geometries. Their results demonstrate that the non‑monotonic η(Λ) behavior is robust across a broad parameter space, persisting even in moderately dense and confined suspensions, though its magnitude diminishes with increasing concentration and swelling.
In conclusion, the paper provides a comprehensive numerical validation of the theoretical prediction that microscopic vesicle dynamics directly dictate macroscopic suspension viscosity. By systematically exploring concentration, membrane rigidity, swelling, and confinement, it clarifies under which conditions the characteristic viscosity minimum appears and why some studies may have missed it. The findings have practical implications for designing complex fluids (e.g., blood analogues, drug‑delivery carriers) where controlling the viscosity through vesicle properties is desirable.
Comments & Academic Discussion
Loading comments...
Leave a Comment