Modelling cytoskeletal traffic: an interplay between passive diffusion and active transport
We introduce the totally asymmetric exclusion process with Langmuir kinetics (TASEP-LK) on a network as a microscopic model for active motor protein transport on the cytoskeleton, immersed in the diffusive cytoplasm. We discuss how the interplay between active transport along a network and infinite diffusion in a bulk reservoir leads to a heterogeneous matter distribution on various scales. We find three regimes for steady state transport, corresponding to the scale of the network, of individual segments or local to sites. At low exchange rates strong density heterogeneities develop between different segments in the network. In this regime one has to consider the topological complexity of the whole network to describe transport. In contrast, at moderate exchange rates the transport through the network decouples, and the physics is determined by single segments and the local topology. At last, for very high exchange rates the homogeneous Langmuir process dominates the stationary state. We introduce effective rate diagrams for the network to identify these different regimes. Based on this method we develop an intuitive but generic picture of how the stationary state of excluded volume processes on complex networks can be understood in terms of the single-segment phase diagram.
💡 Research Summary
The paper presents a theoretical framework for describing the collective transport of motor proteins along the cytoskeletal network while interacting with a diffusive cytoplasmic reservoir. The authors extend the classic totally asymmetric exclusion process (TASEP) by incorporating Langmuir kinetics (LK) – i.e., stochastic attachment and detachment of particles to a bulk reservoir – and embed this TASEP‑LK dynamics on a generic graph representing the filament network. Each edge of the graph is a one‑dimensional lattice of identical sites; particles hop unidirectionally subject to hard‑core exclusion, and they may also bind from or unbind to the reservoir with rates ω_on and ω_off. The reservoir is assumed infinite, so its concentration remains constant.
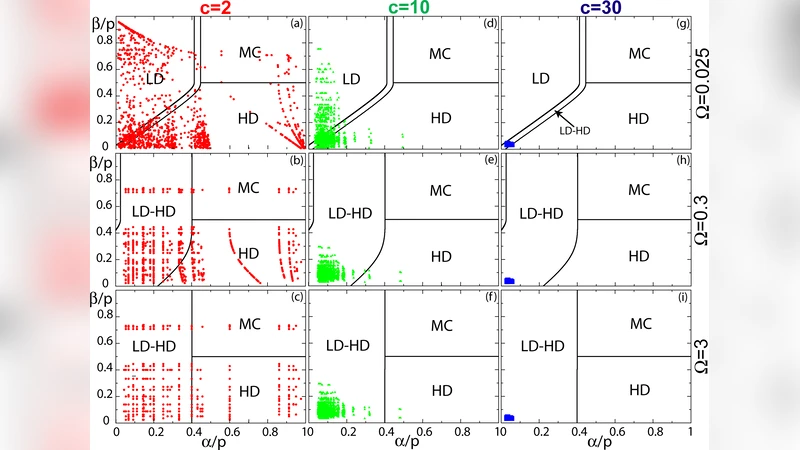
A central analytical tool introduced is the “effective rate diagram”. For any edge i, the effective entry (α_i^eff) and exit (β_i^eff) rates are functions of the local vertex occupancies and the network connectivity. By mapping (α_i^eff, β_i^eff) onto the well‑known phase diagram of a single TASEP‑LK segment (low‑density (LD), high‑density (HD), maximal‑current (MC), and Langmuir‑dominated regimes), the authors can infer the phase of each edge and, consequently, the global stationary state of the whole network.
Three distinct regimes emerge depending on the magnitude of the exchange parameter Ω = ω_on + ω_off (or, equivalently, the ratio of attachment to detachment rates):
-
Low‑exchange regime (Ω ≪ 1).
Attachment/detachment events are rare; transport is dominated by the internal hopping dynamics. Effective entry and exit rates of neighboring edges become strongly coupled, so the density profile of one segment influences many others. Network topology (vertex degree, presence of loops, branching points) strongly shapes the stationary density distribution, leading to pronounced heterogeneities across the network. The system must be treated as a whole; single‑segment analysis is insufficient. -
Intermediate‑exchange regime (Ω ≈ O(1)).
Exchange with the bulk is frequent enough to partially decouple edges. Each edge behaves almost independently, and its phase is determined mainly by the local connectivity (how many edges feed into or out of its end vertices). The global state can be understood by superimposing the single‑segment phase diagram on each edge, with most edges lying near the MC line, indicating efficient transport. This regime represents a balance between active transport along filaments and replenishment from the cytoplasm. -
High‑exchange regime (Ω ≫ 1).
Langmuir kinetics dominate; particles rapidly bind and unbind, erasing the memory of exclusion‑driven jams. All edges converge to the same homogeneous density ρ_eq = ω_on c / (ω_on c + ω_off), where c is the reservoir concentration. The classic TASEP phases disappear, and the system behaves like a well‑mixed Langmuir process.
The authors validate these predictions through mean‑field calculations and extensive Monte‑Carlo simulations on various graph ensembles (regular lattices, Erdős‑Rényi random graphs, scale‑free networks). They demonstrate that the phase boundaries predicted by the effective rate diagram match simulation results across a wide range of system sizes (N ≈ 10^2–10^4) and average degrees. Moreover, they show how the three regimes manifest in observable quantities such as total current, density variance, and correlation length.
Biologically, the three regimes correspond to different cellular strategies. In the low‑exchange limit, transport is highly directed and concentrated along specific “highway” filaments, which could be advantageous for rapid delivery of cargo to particular organelles. In the intermediate regime, the cell achieves a compromise: cargo is efficiently moved along filaments while the cytoplasmic pool supplies and removes particles, allowing flexible redistribution. In the high‑exchange limit, the system behaves like a diffusion‑dominated environment, ensuring uniform distribution of soluble factors.
The paper concludes that the effective‑rate diagram provides a powerful, intuitive method to reduce the complexity of excluded‑volume transport on arbitrary networks to the well‑understood physics of a single TASEP‑LK segment. This reduction opens the door to quantitative modeling of intracellular transport on realistic cytoskeletal architectures extracted from microscopy data, and suggests extensions such as bidirectional motors, dynamic filament remodeling, and heterogeneous attachment/detachment rates.
Comments & Academic Discussion
Loading comments...
Leave a Comment