Neutral and Stable Equilibria of Genetic Systems and The Hardy-Weinberg Principle: Limitations of the Chi-Square Test and Advantages of Auto-Correlation Functions of Allele Frequencies
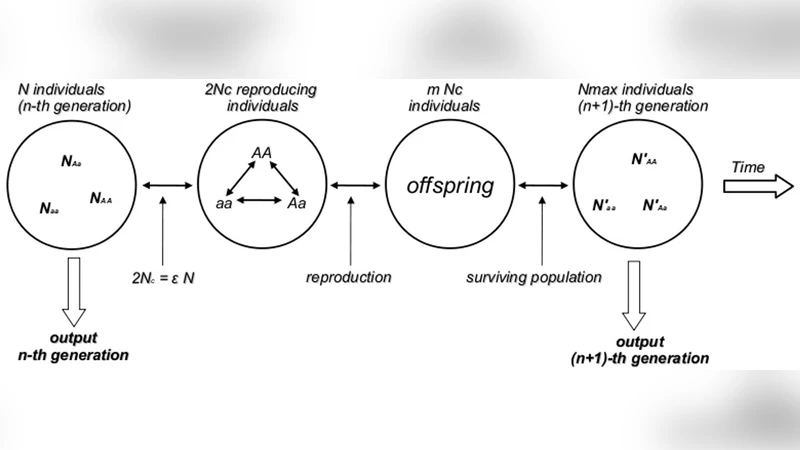
Since the foundations of Population Genetics the notion of genetic equilibrium (in close analogy to Classical Mechanics) has been associated to the Hardy-Weinberg (HW) Principle and the identification of equilibrium is currently assumed by stating that the HW axioms are valid if appropriate values of Chi-Square (p<0.05) are observed in experiments. Here we show by numerical experiments with the genetic system of one locus/two alleles that considering large ensembles of populations the Chi-Square test is not decisive and may lead to false negatives in random mating populations and false positives in nonrandom mating populations. As a result we confirm the logical statement that statistical tests can not be used to deduce if the genetic population is under the HW conditions. Furthermore, we show that under the HW conditions populations of any finite size evolve in time according to what can be identified as neutral dynamics to which the very notion of equilibrium is unattainable for any practical purpose. Therefore, under the HW conditions equilibrium properties are not observable. We also show that by relaxing the condition of random mating the dynamics acquires all the characteristics of asymptotic stable equilibrium. As a consequence our results show that the question of equilibrium in genetic systems should be approached in close analogy to non-equilibrium statistical physics and its observability should be focused on dynamical quantities like the typical decay properties of the allelic auto correlation function in time. In this perspective one should abandon the classical notion of genetic equilibrium and its relation to the HW proportions and open investigations in the direction of searching for unifying general principles of population genetic transformations capable to take in consideration these systems in their full complexity.
💡 Research Summary
The paper revisits the long‑standing identification of Hardy‑Weinberg (HW) equilibrium with a mechanical notion of static balance and challenges the common practice of using a single‑point chi‑square (χ²) test (p < 0.05) to declare that a population satisfies HW conditions. Using extensive Monte‑Carlo simulations of a single‑locus, two‑allele system, the authors generate thousands of independent populations under two regimes: (i) strict random mating, which fulfills the classical HW assumptions, and (ii) a biased, non‑random mating scheme that favours homozygotes. For each generation they record allele frequencies (p, q) and genotype counts, compute χ² against the expected HW proportions, and evaluate the p‑value.
The first major finding is that even when the random‑mating assumptions are perfectly met, the χ² test frequently yields both false‑negative (p > 0.05) and false‑positive (p < 0.05) outcomes. The variability stems from finite‑population sampling noise, the choice of initial genotype distribution, and the fact that χ² is a snapshot statistic that ignores temporal dynamics. Consequently, the test cannot decisively confirm or reject HW equilibrium.
The second finding concerns the biased mating scenario. Here χ² almost always signals a deviation from HW (p < 0.05), yet the time series of allele frequencies shows a clear convergence toward a new stationary point. To capture this behaviour the authors compute the autocorrelation function of allele frequency, C(τ) = ⟨p(t)p(t + τ)⟩ − ⟨p⟩². Under random mating C(τ) decays extremely slowly or remains flat, indicating neutral dynamics: the population wanders indefinitely without a well‑defined equilibrium. By contrast, under biased mating C(τ) exhibits exponential decay with a characteristic time τ₀, demonstrating asymptotic stable equilibrium. This dichotomy mirrors concepts from non‑equilibrium statistical physics, where the presence of restoring forces (here, non‑random mating) creates a measurable relaxation time, whereas neutral drift lacks such a timescale.
From these results the authors draw two overarching conclusions. First, the detection of HW equilibrium should shift from static goodness‑of‑fit tests to dynamical diagnostics such as autocorrelation decay, spectral density analysis, or Markov transition matrices that quantify how quickly allele frequencies lose memory of their initial state. Second, because real populations are finite and never achieve perfect random mating, the classical HW “equilibrium” is, in practice, an unobservable idealization. In finite random‑mating populations the system is perpetually in a neutral drift regime, making the notion of a static equilibrium meaningless for empirical work.
The paper therefore advocates a paradigm shift: population genetics ought to adopt the language and tools of non‑equilibrium statistical physics, focusing on measurable dynamical quantities rather than on static genotype proportions. By doing so, researchers can develop unified principles that accommodate the full complexity of real genetic systems—including selection, migration, mutation, and assortative mating—without relying on the misleading inference that a low χ² automatically guarantees Hardy‑Weinberg equilibrium.
Comments & Academic Discussion
Loading comments...
Leave a Comment