Cooperative Lattice Dynamics and Anomalous Fluctuations of Microtubules
Microtubules have been in biophysical focus for several decades. Yet the confusing and mutually contradicting results regarding their elasticity and fluctuations have shed some doubts on their present understanding. In this paper we expose the empirical evidence for the existence of discrete GDP-tubulin fluctuations between a curved and a straight configuration at room temperature as well as for conformational tubulin cooperativity. Guided by a number of experimental findings, we build the case for a novel microtubule model, with the principal result that microtubules can spontaneously form micron size cooperative helical states with unique elastic and dynamic features. The polymorphic dynamics of the microtubule lattice resulting from the tubulin bistability quantitatively explains several experimental puzzles including anomalous scaling of dynamic fluctuations of grafted microtubules, their apparent length-stiffness relation and their remarkably curved-helical appearance in general. We point out that tubulin dimers’s multistability and its cooperative switching could participate in important cellular processes, and could in particular lead to efficient mechanochemical signalling along single microtubules.
💡 Research Summary
The paper tackles a long‑standing paradox in microtubule (MT) mechanics: conventional models—either a simple Euler‑Bernoulli beam or the “soft‑shear” (Timoshenko) composite description—cannot simultaneously account for the wide scatter of measured persistence lengths, the observed length‑dependent stiffness, the anomalous L³ scaling of relaxation times, and the frequent appearance of permanent curvature or helical shapes in taxol‑stabilized MTs.
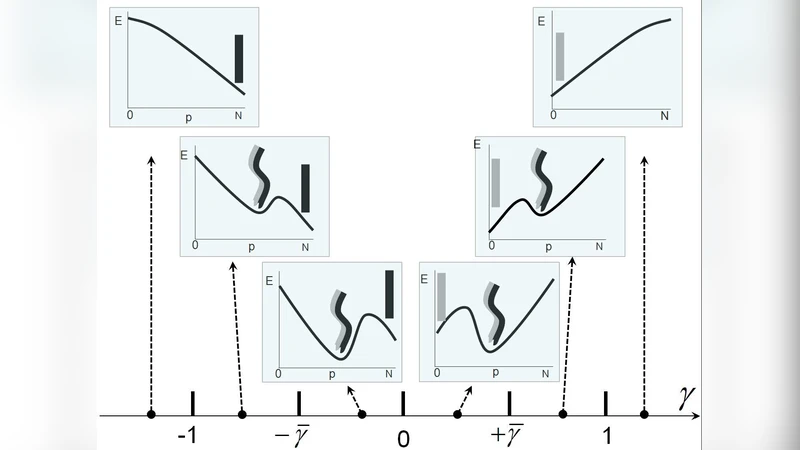
To resolve these contradictions the authors propose a “polymorphic” MT model based on two experimentally motivated assumptions: (I) a taxol‑bound GDP‑tubulin dimer is bistable, fluctuating between a straight conformation (curvature κ≈0) and a weakly curved state (κ≈1/250 nm⁻¹) on experimental time scales; (II) neighboring dimers along a protofilament interact cooperatively, favoring alignment of their conformational states.
Mathematically the MT lattice is mapped onto a one‑dimensional Ising‑type chain where each protofilament segment carries a binary variable σ = ±1 (straight/curved). Nearest‑neighbor coupling J encodes the cooperative interaction, while an effective field h represents the intrinsic bias imposed by the lattice geometry (the native 13‑protofilament twist of +3.4° per subunit). The total energy includes a bending term that couples the σ‑field to the global curvature of the tube.
Solving this model (mean‑field and Monte‑Carlo simulations) yields several key predictions:
-
Spontaneous cooperative helices – Above a critical length Lc, domains of aligned σ‑states nucleate, producing a global helical deformation of the MT with a well‑defined pitch and radius. The pitch is set by the ratio J/h and the intrinsic lattice twist; typical values match the experimentally observed 15 µm pitch helices.
-
Length‑dependent persistence length – The presence of domain walls reduces the effective bending rigidity B_eff ∝ 1/L, leading to a persistence length l_p = B_eff/kBT that scales linearly with MT length, exactly as reported in grafted‑MT experiments. Short MTs (< 4 µm) remain essentially straight because domain formation is suppressed, reproducing the “plateau” region of stiffness.
-
Anomalous dynamics – The helical domains introduce a low‑energy “zero‑mode” corresponding to a collective rotation of the helix about its axis. For clamped MTs this mode dominates the longest relaxation time, giving τ_max ∝ L³ rather than the L⁴ expected for a simple semiflexible filament. This explains the unusually slow thermal dynamics measured in several optical‑trap and fluorescence studies.
-
Permanent curvature and circular gliding – The model naturally accounts for the persistent sinusoidal, circular, and helical shapes observed when MTs are adsorbed on glass, confined between surfaces, or propelled by surface‑bound kinesin motors. The cooperative bistability allows the tube to lock into a curved state without external forces, consistent with the “frozen‑in” curvature reported by Amos & Amos and by Venier et al.
-
Compatibility with drug effects – Because J and h can be modulated by taxol, MAPs, or other binding agents, the model predicts that pharmacological manipulation will shift the helix pitch, domain size, and effective stiffness—providing a testable route to validate the theory.
Beyond mechanics, the authors suggest that the bistable tubulin lattice could serve as a mechano‑chemical signaling conduit: a local mechanical perturbation (e.g., motor force, filament bending) could trigger a cooperative conformational wave that propagates over microns, thereby coupling mechanical cues to biochemical states along a single MT.
In summary, the paper introduces a unified physical framework that reconciles a host of puzzling experimental observations on microtubules. By invoking cooperative switching of multistable tubulin dimers, it explains the emergence of micron‑scale helical superstructures, the length‑dependent elastic response, and the anomalous relaxation dynamics, while also opening new perspectives on mechanotransduction within the cytoskeleton.
Comments & Academic Discussion
Loading comments...
Leave a Comment