Bcl-2 inhibits apoptosis by increasing the time-to-death and intrinsic cell-to-cell variations in the mitochondrial pathway of cell death
BH3 mimetics have been proposed as new anticancer therapeutics. They target anti-apoptotic Bcl-2 proteins, up-regulation of which has been implicated in the resistance of many cancer cells, particularly leukemia and lymphoma cells, to apoptosis. Using probabilistic computational modeling of the mitochondrial pathway of apoptosis, verified by single-cell experimental observations, we develop a model of Bcl-2 inhibition of apoptosis. Our results clarify how Bcl-2 imparts its anti-apoptotic role by increasing the time-to-death and cell-to-cell variability. We also show that although the commitment to death is highly impacted by differences in protein levels at the time of stimulation, inherent stochastic fluctuations in apoptotic signaling are sufficient to induce cell-to-cell variability and to allow single cells to escape death. This study suggests that intrinsic cell-to-cell stochastic variability in apoptotic signaling is sufficient to cause fractional killing of cancer cells after exposure to BH3 mimetics. This is an unanticipated facet of cancer chemoresistance.
💡 Research Summary
This paper investigates how the anti‑apoptotic protein Bcl‑2 confers resistance to cell death by modulating both the timing of mitochondrial outer‑membrane permeabilization (MOMP) and the intrinsic cell‑to‑cell variability of the intrinsic apoptosis pathway. The authors combine a probabilistic computational model of the mitochondrial apoptosis cascade with single‑cell experimental measurements to dissect the mechanistic underpinnings of Bcl‑2–mediated chemoresistance, particularly in the context of BH3‑mimetic drugs that target Bcl‑2 family proteins.
Model Development and Validation
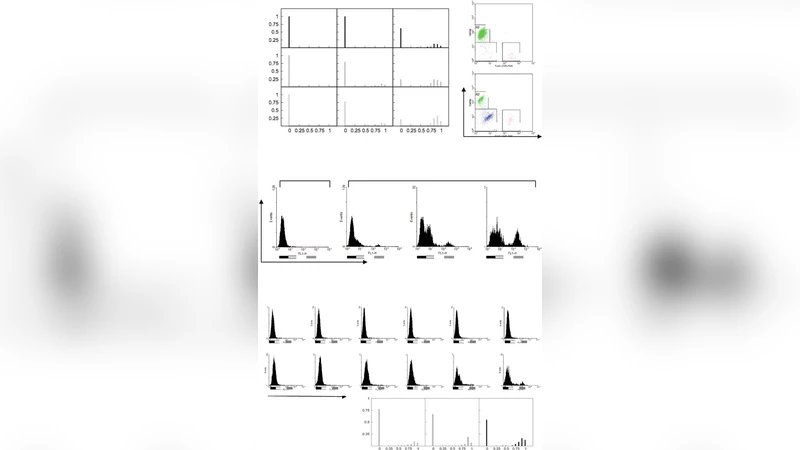
Starting from a deterministic ordinary‑differential‑equation (ODE) framework that describes the interactions among pro‑apoptotic proteins (Bax, Bak, tBid), anti‑apoptotic Bcl‑2, cytochrome c release, Apaf‑1, and downstream caspases, the authors introduce stochasticity by converting each reaction into a Poisson process. They implement Gillespie’s stochastic simulation algorithm to generate single‑cell trajectories, allowing them to capture the random timing of key events such as Bax activation, MOMP, and caspase activation. Model parameters are calibrated using literature values and experimental data (Western blots, flow cytometry) from human leukemia cell lines (e.g., HL‑60) and engineered Bcl‑2‑overexpressing variants.
Experimental Approach
To validate the model, the authors use live‑cell imaging to monitor MOMP in individual cells after a controlled tBid stimulus (delivered by electroporation). They employ TMRE (a mitochondrial membrane potential dye) and a cytochrome c‑GFP fusion to record the loss of membrane potential and the release of cytochrome c, respectively. Time‑to‑death is defined as the interval between tBid introduction and irreversible loss of TMRE signal. Parallel experiments are performed with the BH3‑mimetic ABT‑737, alone or in combination with Bcl‑2 overexpression, to assess drug‑induced killing efficiency.
Key Findings
-
Bcl‑2 Extends Time‑to‑Death
Simulations predict that increasing Bcl‑2 levels raise the threshold of activated Bax/Bak required for MOMP. Experimentally, Bcl‑2‑overexpressing cells display a 2–3‑fold delay in MOMP compared with control cells under identical tBid stimulation, confirming the model’s prediction. -
Intrinsic Stochastic Variability Drives Fractional Killing
Even when initial protein concentrations are held constant, stochastic simulations generate a broad distribution of death times, with a non‑negligible fraction of trajectories never reaching MOMP. Single‑cell imaging mirrors this behavior: a subset of cells never loses TMRE signal despite exposure to the same stimulus. The probability of survival correlates positively with Bcl‑2 abundance, indicating that Bcl‑2 amplifies the impact of molecular noise. -
BH3‑Mimetics Reduce but Do Not Eliminate Variability
Treatment with ABT‑737 dramatically increases the overall killing fraction, yet a residual population of “escapees” persists, especially in cells with high Bcl‑2 expression. The model shows that while the drug lowers the Bax activation threshold, stochastic fluctuations still allow some cells to remain below the critical level needed for irreversible MOMP. -
Implications for Chemoresistance
The authors argue that fractional killing is not solely a consequence of heterogeneous drug penetration or cell‑cycle phase differences; rather, it can arise from the inherent stochastic nature of the apoptosis signaling network, which Bcl‑2 exploits. Consequently, therapeutic regimens that target Bcl‑2 alone may be insufficient to achieve complete tumor eradication.
Broader Significance
The study introduces a quantitative framework that integrates stochastic modeling with high‑resolution single‑cell data, offering a powerful tool to predict how variations in protein expression and molecular noise influence therapeutic outcomes. It suggests that future anti‑cancer strategies should aim not only to inhibit anti‑apoptotic proteins but also to dampen the underlying stochastic fluctuations—potentially by combining BH3‑mimetics with agents that either sensitize Bax/Bak activation or amplify downstream caspase signaling.
Conclusion
Bcl‑2 confers resistance by both lengthening the latency before MOMP and by magnifying intrinsic cell‑to‑cell variability in the mitochondrial apoptosis pathway. This dual effect creates a population of cells that can survive BH3‑mimetic treatment, providing a mechanistic explanation for the observed fractional killing in cancer therapy. The findings underscore the necessity of accounting for stochastic cellular behavior when designing and evaluating apoptosis‑targeted drugs, paving the way for more robust combination therapies that can overcome the stochastic barrier to complete tumor cell elimination.
Comments & Academic Discussion
Loading comments...
Leave a Comment