A waiting time problem arising from the study of multi-stage carcinogenesis
We consider the population genetics problem: how long does it take before some member of the population has $m$ specified mutations? The case $m=2$ is relevant to onset of cancer due to the inactivation of both copies of a tumor suppressor gene. Mode…
Authors: ** R. Durrett, D. Schmidt, J. Schweinsberg **
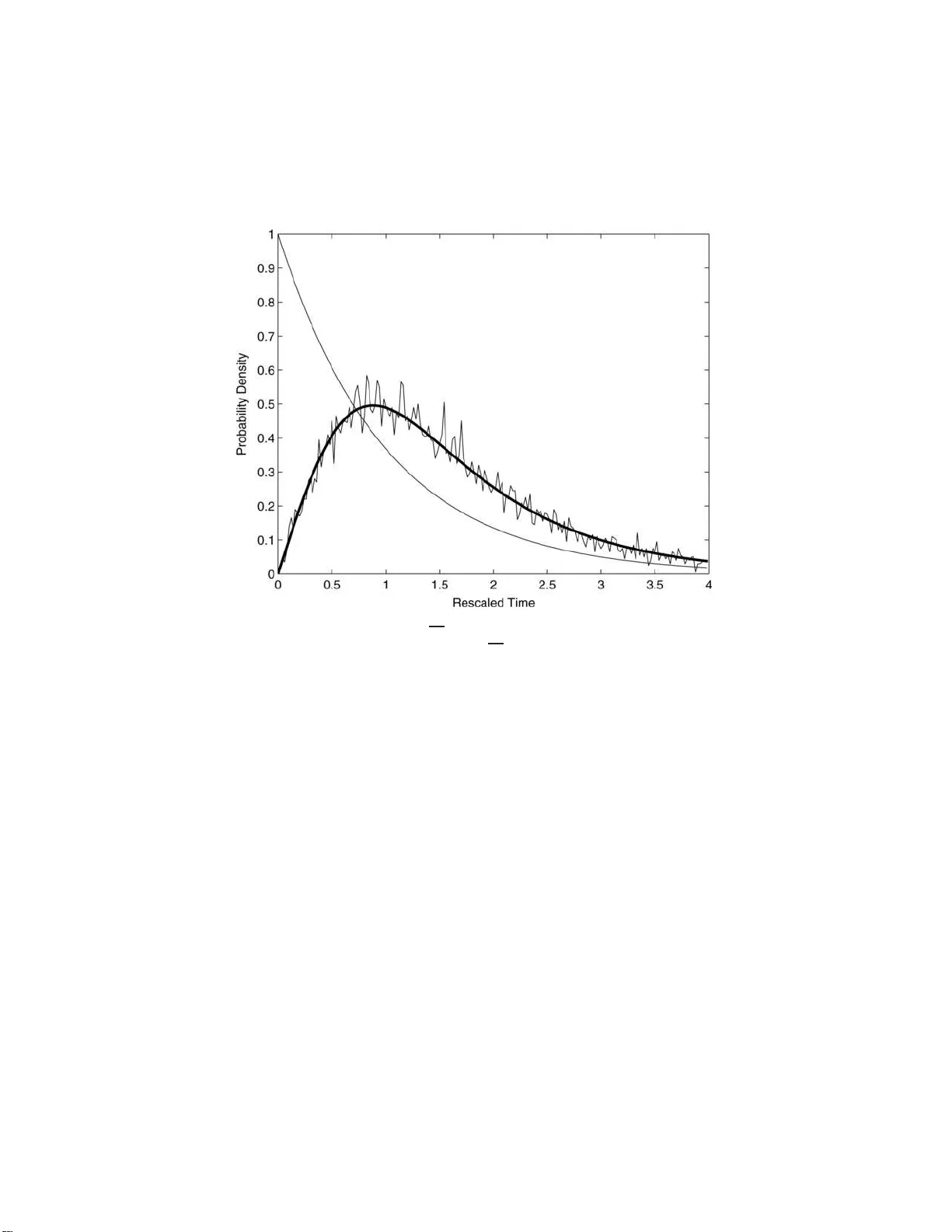
The Annals of Applie d Pr obabil ity 2009, V ol. 19, N o. 2, 676– 718 DOI: 10.1214 /08-AAP559 c Institute of Mathematical Statistics , 2 009 A W AITING TIME PR OBL EM ARISING FR OM THE STUD Y OF MUL TI-ST AGE CAR CINOGENESIS By Rick Durrett, 1 Deena Schmidt 1 , 2 and Jason Schweinsb e rg 3 Cornel l University, University o f Minnesota and University of California, San Die go W e consider the p opulation genetics problem: how l ong do es it take before some mem b er of the p opu lation has m specified m ut a- tions? The case m = 2 is relev ant to on set of cancer due to th e inac- tiv ation of b oth copies of a tumor supp ressor gene. Models for larger m are needed for colon cancer and other diseases where a sequen ce of m utations leads to cells with uncontro lled gro wth. 1. In tro d uction. It has long b een kn own that cancer is the end resu lt of sev eral m u tations that disrupt normal ce ll division. Armitage and Doll [ 1 ] did a statistical analysis of the age of onset of sev eral cancers and fit p o wer la ws to estimate the num b er of m u tations. Knudson [ 15 ] disco vered that the incidence of retinoblastoma (cancer of the retina) gro ws as a linear function of time in the group of c hildren who hav e m u ltiple cancers in b oth ey es, but as a slo wer quadr atic f unction in children wh o only ha v e one cancer. Based on this, Knudson prop osed the concept of a tumor supp ressor gene. Later it w as confirmed that in the first group of c hildren, one cop y is already inactiv ated at birth , wh ile in the second group b oth copies m u st b e m u tated b efore cancer occur s. S ince that time, ab out 30 tumor supp r essor genes ha ve b een id entified. They hav e the prop erty that inactiv ating the first cop y do es not cause a change, w hile inactiv ating the second increases the cells’ n et repro du ctiv e rate, whic h is a step to w ard cancer. There is n ow considerable evidence that colon cancer is the end result of sev eral m utations. Th e earliest evidence w as statistical. Lueb eck and Mo ol- Received July 2007; revised July 2008. 1 Supp orted in part by NSF Grant DMS-02-02935 from th e probabilit y program and NSF/NIGMS Grant DMS-02-01037. 2 Supp orted by an NS F graduate fello w. 3 Supp orted in part by NSF Gran ts D MS-05-04882 and DMS-08-05472. AMS 2000 subje ct classific ations. Primary 60J99; secondary 92C50, 92D25, 60J85. Key wor ds and phr ases. Multi-stage carcinogenesis, wa iting times, Mora n model, branching process, W right– Fisher diffusion. This is an electronic repr int of the or iginal ar ticle publishe d b y the Institute of Mathematical Statistics in The A n n als of Applie d Pr ob ability , 2009, V o l. 19, No. 2, 67 6–71 8 . This reprint differs fro m the or iginal in pa gination and typo graphic detail. 1 2 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG ga v ak ar [ 18 ] fit a four-stage mo del to the age-sp ecific incidence of colorectal cancers in the Surv eillance, Epidemiology , and End Results registry , wh ich co v er 10 p ercent of the US p opu lation. Calabrese et al. [ 5 ] examined 1022 colorecta l cancers sampled fr om nine large r egional hosp itals in southeastern Finland. They found supp ort for a mo del w ith fiv e or six oncogenic m u ta- tions in individu als with hereditary r isk factors and seven or eigh t m utations in patien ts without. Ov er the last decade, a num b er of studies ha ve b een carried out to iden tify the molecular pathw a ys in volv ed in the develo p men t of colorectal cancer. S ee Jones et al. [ 14 ] for a recen t r ep ort. The pro cess is initiated when a single colorecta l cell acquires mutatio ns inactiv ating the ACP/ β -catenin pathw a y . This results in the gro wth of small b enign tumor (adenoma). Su bsequent m u tations in a short list of other pathw a ys transform the aden oma into a malig nan t tum or (carcinoma), and lead to metastasis, the abilit y of the cancer to spread to other organs. In th is p ap er, w e pr op ose a simple mathematical mo d el f or cancer de- v elopment in whic h cancer o ccurs when one cell accum ulates m m u tations. Consider a p opulation of fixed size N . Readers who are u sed to the study of th e genetics of diploid organisms ma y ha ve exp ected to see 2 N here, but our concern is for a collection of N cells. W e choose a m o del in whic h the n u m b er of cells is fixed b ecause organs in the b o dy are t ypically of constant size. W e assume that the p opulation ev olves according to the Moran mo del, whic h was fir st p rop osed b y Moran [ 19 ]. That is, eac h individu al liv es for an exp onenti ally distrib uted amount of time w ith mean one, and then is replaced by a new ind ivid ual whose p aren t is c hosen at random from th e N individuals in the p opulation (including the one b eing replaced). F or more on this mod el, see Section 3.4 of [ 11 ]. In our mo del, eac h in d ividual has a t yp e 0 ≤ j ≤ m . Initially , all individ- uals ha ve t yp e 0. In the u sual p opulation genetics mo del, m utations only o ccur at replacemen t ev ents. W e assume instead that t yp es are clonally in- herited, that is, every individual has the same typ e as its paren t. How ev er, thinking of a collection of cells that ma y acquire mutati ons due to r adiation or other en vironmental factors, we will sup p ose that du ring their lifetimes, individuals of t yp e j − 1 m u tate to typ e j at rate u j . W e call such a mu- tation a typ e j mutation . Let X j ( t ) b e the n um b er of t yp e j in dividuals at time t . F or ea c h p ositiv e in teger m , let τ m = in f { t : X m ( t ) > 0 } b e the fi rst time at wh ic h there is an in d ividual in the p opulation of t yp e m . Clearly , τ 1 has the exp onen tial distribu tion with rate N u 1 . Our goa l is to compute the asymptotic distribution of τ m for m ≥ 2 as N → ∞ . W e b egin b y consider in g the case m = 2 and discu s sing previous work. Sc h inazi [ 21 , 22 ] has considered related questions. In the fir s t pap er, he computes the pr obabilit y that in a br an ching pro cess wher e individuals ha ve t wo offspr ing with probabilit y p and zero with probabilit y 1 − p , a m u tation W AITIN G TIMES F OR MUT A TIONS 3 will arise b efore the pro cess dies out. In the second pap er, he uses this to in vestig ate the probabilit y of a t yp e 2 mutatio n when t yp e 0 cells divide a fixed n u m b er of times w ith the p ossib ilit y of m u tating to a t yp e 1 cell that b egins a binary br anc hin g p ro cess. More relev an t to our inv estigat ion is the w ork of Komaro v a, S engupta and No wak [ 17 ], Iwa sa, Mic hor an d No w ak [ 13 ] and Iw asa et al. [ 12 ]. Their analysis b egins with the observ ation that w hile the num b er of m utant in- dividuals is o ( N ), we can approximate the num b er of cells with m utations b y a branc hin g pro cess in w hic h eac h individu al giv es birth at rate one and dies at rate one. Let Z denote the tota l pr ogen y of such a branching pro cess. Since th e em b edd ed discrete time Mark o v c hain is a simple rand om w alk, w e hav e (see, e.g., p age 197 in [ 7 ]) P ( Z > n ) = 2 − 2 n 2 n n ∼ 1 √ π n . If we ignore interference b etw een successiv e new type 1 mutati ons, then their total p rogen y Z 1 , Z 2 , . . . are i.i.d. v ariables in the domain of attrac tion of a stable law with in dex 1 / 2, so max i ≤ M Z i and Z 1 + · · · + Z M will b e O ( M 2 ). Therefore, w e exp ect to see ou r fir st t yp e 2 m u tation in the family of th e M th t yp e 1 mutatio n , where M = O (1 / √ u 2 ). S tandard results for simple random wal k imply that the largest of our fi rst M families will hav e O ( M ) t yp e 1 ind ividuals aliv e at th e same time, so for the b ranc h ing pro cess appro ximation to hold, w e n eed 1 / √ u 2 ≪ N , w here h er e and thr ou gh ou t the p ap er, f ( N ) ≪ g ( N ) means that f ( N ) /g ( N ) → 0 as N → ∞ . Type 1 m u tations o ccur at rate N u 1 , so a typ e 2 mutation will first o ccur at a time of order 1 / N u 1 √ u 2 . As long as the branching pro cess approximati on is accurate, the amount of time w e hav e to wait for a t yp e 1 m utation that will ha ve a t yp e 2 ind ividual as a descendan t will b e approximat ely exp onential, since mutatio ns o ccur at times of a Po iss on pro cess with rate N u 1 and the type 1 muta tions that lead to a type 2 are a th inning of that pr o cess in w h ic h p oin ts are k ept with probabilit y ∼ √ u 2 , whic h is O (1 / M ) , where here and thr oughout the pap er, f ( N ) ∼ g ( N ) means that f ( N ) /g ( N ) → 1 as N → ∞ . The duration of the longest of M t yp e 1 families is O ( M ), s o the time b et we en when the t yp e 1 m utation o ccurs and when the type 2 descendan t app ears is O (1 / √ u 2 ). This will b e negligible in comparison to 1 / N u 1 √ u 2 as long as N u 1 ≪ 1, so the w aiting time for the fir st t y p e 2 individu al will also b e appro ximately exp onentia l. T h is leads to a result s tated on pages 231–232 of No wak’s b ook [ 20 ] on E volutionary Dynamics. If 1 / √ u 2 ≪ N ≪ 1 /u 1 , th en P ( τ 2 ≤ t ) ≈ 1 − exp( − N u 1 √ u 2 t ) . (1.1) Figure 1 sho ws the distribution of τ 2 · N u 1 √ u 2 in 10,000 simulatio n s of the Moran mod el when N = 10 3 and u 1 = u 2 = 10 − 4 . Here, N u 1 = 0 . 1 and 4 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG Fig. 1. Distribution of τ 2 · N u 1 √ u 2 = 1000 in 10,000 simul ations when N = 10 3 and u 1 = u 2 = 10 − 4 . N u 1 = 0 . 1 and N √ u 2 = 10 , so as ( 1.1 ) pr e dicts the sc ale d wai ting time is appr oximately exp onential. N √ u 2 = 10, so as the last r esult p redicts, the scaled w aiting time is approx- imately exp onent ial. W e do n ot refer to the result giv en in ( 1.1 ) as a theorem b ecause their argu- men t is n ot complete ly rigorous. F or example, the au th ors use the b r anc hin g pro cess appro xim ation w ithout pro vin g it is v alid. Ho wev er, this is a minor quibble, since as the reader will see in S ection 2 , it is straigh tforwa rd to fi ll in the missing details and establish the follo wing more general r esult. Theorem 1. Supp ose that N u 1 → λ ∈ [0 , ∞ ) , u 2 → 0 and N √ u 2 → ∞ as N → ∞ . Then τ 2 · N u 1 √ u 2 c onver ges to a limit that has density function f 2 ( t ) = h ( t ) exp − Z t 0 h ( s ) ds wher e h ( s ) = 1 − e − 2 s/λ 1 + e − 2 s/λ , if λ > 0 and f 2 ( t ) = e − t if λ = 0 . Here, h ( t ) is the h azard function, that is, if w e let F 2 ( t ) = exp ( − R t 0 h ( s ) ds ) b e the tail of the distribution, then h ( t ) = f 2 ( t ) /F 2 ( t ). Figure 2 sho w s the distribution of τ 2 · N u 1 √ u 2 in 10,000 sim ulations of the Moran mo d el when W AITIN G TIMES F OR MUT A TIONS 5 Fig. 2. Distribution of τ 2 · N u 1 √ u 2 = 1000 in 10,000 simul ations when N = 10 3 , u 1 = 10 − 3 and u 2 = 10 − 4 . N u 1 = 1 and N √ u 2 = 0 . 1 , so the limit is not exp onential, but is fit wel l by the r esult in The or em 1 . N = 10 3 , u 1 = 10 − 3 and u 2 = 10 − 4 . N u 1 = 1 so the limit is not exp on ential, but Theorem 1 giv es a goo d fi t to the observ ed d istribution. Before turn ing to the case of m mutat ions, we should clarify one p oin t. In our mo del, mutations o ccur during the lifetime of an ind ividual, but in the follo wing discuss ion, w e will coun t bir ths to estimate the probabilit y a desired mutatio n will o ccur. This migh t seem to only b e appropr iate if muta - tions o ccur at birth. Ho wev er, since eac h individ u al liv es for an exp onen tial amoun t of time with mean 1, the num b er of “man-h ou r s” R T 0 0 X 1 ( s ) ds b e- fore the family dies out at time T 0 is roughly the same as the n u m b er of births. In any case, the follo wing discuss ion is only a heur istic that helps explain the answ er, but do es not dir ectly en ter into its pro of. T o extend the analysis to the m -stage w aiting time problem, supp ose M distinct t yp e 1 mutations h av e app eared. If the family sizes of th ese M m u tations can b e mo deled by indep enden t branching pro cesses, the total n u m b er of offspr in g of t yp e 1 in d ividuals will b e O ( M 2 ). Because eac h type 1 individu al m u tates to t yp e 2 at r ate u 2 , there will b e O ( M 2 u 2 ) mutat ions that p ro duce type 2 individuals. T he total progen y of these individuals will consist of O ( M 4 u 2 2 ) type 2 individu als. W e can exp ect to see our first t yp e 6 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG 3 individu al when M 4 u 2 2 = O (1 /u 3 ) or M = O ( u − 1 / 2 2 u − 1 / 4 3 ). Thus, for the branc hing p ro cess app ro ximation to h old, we need u − 1 / 2 2 u − 1 / 4 3 ≪ N . Since t yp e 1 m u tations o ccur at rate N u 1 , the exp ected w aiting time will b e of order 1 / N u 1 u 1 / 2 2 u 1 / 4 3 . T o help deve lop a go o d menta l picture, it is instructiv e to consider th e n u merical example in which N = 10 5 , u 1 = 10 − 6 , u 2 = 10 − 5 and u 3 = 10 − 4 . By the r easoning ab o ve , w e will fi rst see a t yp e 3 mutation when the num b er of t yp e 2’s is of order 100 = 1 / √ u 3 , since in this case there will b e of ord er 10,000 = 1 /u 3 t yp e 2 births b efore the family d ies out. T o h a ve a typ e 2 family reac h size 100, we will need 100 mutatio n s fr om t yp e 1 to t yp e 2, and for this we will need of order 100 /u 2 = 10 7 t yp e 1 births, which will in turn o ccur if the type 1 family reac hes size of ord er 10 7 / 2 ≈ 3162. Note th at X 2 ( t ) ≪ X 1 ( t ) and w ithin the time that the large t yp e 1 family exists, 100’s of t yp e 2 families will b e s tarted and die out. This difference in the time and size scales for the pro cesses X i ( t ) is a complicating f actor in th e pro of, but ultimately it also allo ws us to separate the t yp e 1’s from typ es 2 to m and use induction. Extrap olating the calculat ion ab ov e to m stages, w e let r j,m = u 1 / 2 j +1 u 1 / 4 j +2 · · · u 1 / 2 m − j m (1.2) for 1 ≤ j < m , an d set r m,m = 1 and r 0 ,m = u 1 r 1 ,m . Let q j,m b e the probabilit y a t yp e j ind ividual giv es r ise to a t yp e m descend ant. W e will show that q j,m ∼ r j,m , s o w e will need of order 1 /r j,m m u tations to type j b efore time τ m . Theorem 2. Fix an inte g e r m ≥ 2 . Supp ose that: (i) N u 1 → 0 . (ii) F or j = 1 , . . . , m − 1 , ther e i s a c onsta nt b j > 0 such that u j +1 /u j > b j for al l N . (iii) Ther e is an a > 0 so that N a u m → 0 . (iv) N r 1 ,m → ∞ . Then for al l t > 0 , lim N →∞ P ( τ m > t/ N r 0 ,m ) = exp ( − t ) . (1.3) As discussed ab o ve , condition (iv) whic h sa ys 1 /r 1 ,m ≪ N is needed for the branc hin g pr o cess assu mption to b e v alid, and condition (i) is needed for the wa iting time to b e exp onentia l, b ecause if (i) fails then th e time b et w een the type 1 muta tion that will hav e a t yp e m descendant and the W AITIN G TIMES F OR MUT A TIONS 7 birth of the t yp e m descendan t cannot b e neglected. If u j = µ for all j , (ii) is trivial. In this case r 1 ,m = µ a ( m ) , wh ere a ( m ) = 1 − 2 − ( m − 1) . Conditions (i) and (iv) b ecome N − 1 /a ( m ) ≪ µ ≪ N − 1 , and w hen condition (i) is satisfied, (iii) holds. Conditions (ii) and (iii) are tec hnicalities that allo w u s to pro ve the result without having to supp ose that u j ≡ µ , which would not b e natu- ral in mo d eling cancer. In the presence of (ii), condition (iii) ensures that max j ≤ m u j ≪ N − a for s ome a > 0. This is natural b ecause ev en in the late stages of pr ogression to cancer, the p er cell division m utation probabilities are small. Condition (ii) is m otiv ated by the fact that in most cancers we exp ect u j to b e increasing in j . T he simple extension of this giv en in (ii) is u s eful so that we do not rule ou t some in teresting sp ecial cases. In mo deling the tumor suppressor genes men tioned earlier, it is natural to take u 1 = 2 µ and u 2 = µ , that is, at the fir s t stage a m u tation can kn o c k out one of th e t wo copies of the ge ne, b ut after this occurs, there is only one copy sub ject to m u tation. A case with u 1 /u 2 = 30 o ccur s in Durr ett and Schmidt’s stu dy of regulatory sequence ev olution [ 9 ]. Condition (iv) ensures that an individual of t yp e m will app ear b efore any t yp e 1 m utation ac hieve s fixation. In the case m = 2, Iw asa et al. [ 13 ] called this sto chastic tunneling . A giv en t yp e 1 mutati on fixates with probabilit y 1 / N and type 1 m utations o ccur at r ate approximat ely N u 1 , so fixation o ccurs b efore a type m individu al app ears if N r 1 ,m → 0, and then once a t yp e 1 muta tion fixates, the problem redu ces to the problem of wait ing for m − 1 additional mutations. In the b ord erline case considered in the next result, either a t yp e m ind ividual could app ear b efore fixation, or a t yp e m mutat ion could b e ac h iev ed through the fixation of t yp e 1 individ uals follo w ed by the generation of an individual w ith m − 1 additional m utations. Theorem 3. Fix an inte ger m ≥ 2 . Assume c onditions (i), (ii) and (iii) fr om The or em 2 hold. If ( N r 1 ,m ) 2 → γ > 0 , and we let α = ∞ X k =1 γ k ( k − 1)!( k − 1)! ∞ X k =1 γ k k !( k − 1)! > 1 , (1.4) then for al l t > 0 , lim N →∞ P ( u 1 τ m > t ) = exp( − αt ) . Figure 3 sh ows the d istribution of u 1 τ 2 in 10,000 sim u lations of the Moran mo del when N = 10 3 , u 1 = 10 − 4 and u 2 = 10 − 6 . N u 1 = 0 . 1 and N √ u 2 = 1, so the assumptions of Theorem 3 hold with γ = 1. Numerically ev aluating the constant giv es α = 1 . 433 and as the fi gure shows the exp onential with this rate giv es a reasonable fi t to the simulat ed data. 8 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG Theorem 3 will b e p r o ved b y redu cing the general case to a t wo-t yp e mo d el with ¯ u 1 = u 1 and ¯ u 2 = u 2 q 2 ,m ∼ r 2 1 ,m . W e will show that it suffices to do calculatio ns for a mo del in which type 1 mutat ions are not allo w ed when th e n u m b er of t yp e 1 individuals X 1 ( t ) is p ositive . In this case, if w e start with X 1 (0) = N ε then N − 1 X 1 ( N t ) → Z t where Z t is the W righ t–Fisher diffusion pro cess with infinitesimal generato r x (1 − x ) d 2 /dx 2 . When X 1 ( N t ) = N x , m u tations to t yp e 2 th at eve n tu ally lead to a t yp e m individual o ccur at rate appro ximately N · N x · u 2 q 2 ,m ∼ N 2 r 2 1 ,m x → γ x, so, if w e let u ( x ) b e the probabilit y that th e p ro cess Z t hits 0 b efore r eac hing 1 or generating a t yp e m m u tation, then u ( x ) satisfies x (1 − x ) u ′′ ( x ) − γ xu ( x ) = 0 , u (0) = 1 , u (1) = 0 . (1.5) The constan t α = lim ε → 0 (1 − u ( ε )) /ε . Its relev ance for the p roblem is that starting from a single typ e 1 individual, the probability of reac hin g N or generating a typ e m m u tation is ∼ α/ N . S ince mutati ons to typ e 1 o ccur at rate ∼ N u 1 , the waiting time is roughly exp onential with rate u 1 α . Fig. 3. Distribution of u 1 τ 2 when N = 10 3 , u 1 = 10 − 4 and u 2 = 10 − 6 . N u 1 = 0 . 1 and N √ u 2 = 1 so we ar e i n the r e gime c over e d by The or em 3 . The c onstan t γ = 1 so α = 1 . 433 . As the gr aph shows the exp onential distribution with r ate α gives a r e asonbly go o d fit to the simulate d data. W AITIN G TIMES F OR MUT A TIONS 9 One can chec k (see Lemma 6.9 b elow) that ( 1.5 ) can b e solv ed b y the follo wing p o wer ser ies around x = 1: u ( x ) = c ∞ X k =1 γ k k !( k − 1)! (1 − x ) k . (1.6) Pic king c so that u (0) = 1, it follo ws that α has the form giv en in ( 1.4 ). Another appr oac h to solving ( 1.5 ) is to use the F eynman–Kac form ula; see form u la (3.19 .5b) on page 225 of [ 4 ]. W e do not d iscuss in this pap er the case N u 1 → ∞ . W e instead refer th e reader to [ 23 ], wh ere asymptotic results in this regime are obtained in th e sp ecial case when u j = µ for all j . The rest of this p ap er is organized as follo ws. In Section 2 , w e giv e the pro of of T h eorem 1 . In Section 3 , w e collect some results for a tw o-t yp e p opulation mo del that will b e useful later in the p ap er. In Section 4 , we calculate by induction the p robabilit y that a giv en t yp e 1 individu al has a type m descendan t. In Section 5 , we com bine this resu lt with a P oisson appro ximation result of Arratia, Goldstein an d Gordon [ 2 ] to pr o ve Theorem 2 . Theorem 3 is prov ed in Sections 6 and 7 . Through ou t our pro ofs, C denotes a constan t whose v alue is un imp ortant and will change from line to line. 2. Pro of of Theorem 1 . If we let X 1 ( t ) b e the num b er of t y p e 1 ind ivid - uals at time t then P ( τ 2 > t ) = E exp − u 2 Z t 0 X 1 ( s ) ds , (2.1) b ecause at time s , there are X 1 ( s ) individuals eac h exp eriencing type 2 m u - tations at rate u 2 . W e will compare X 1 ( t ) with a contin uous-time branching pro cess with immigration, Y ( t ). When X 1 ( t ) = k , t yp e 1 m utations o ccur at r ate ( N − k ) u 1 , wh ile birth ev en ts in whic h a t yp e 1 individual replaces a t yp e 0 ind ividual o ccur at r ate k ( N − k ) / N , so b efore time τ 2 , we hav e jumps k → k + 1 at rate ( k + N u 1 ) · N − k N , k → k − 1 at rate k · N − k N . In the branc hin g pro cess with immigration, Y ( t ), w e hav e jumps k → k + 1 at rate k + N u 1 , k → k − 1 at rate k. Therefore, up to time τ 2 , the p r o cess { X 1 ( t ) , t ≥ 0 } is a time-c hange of { Y ( t ) , t ≥ 0 } , in wh ic h time run s slow er than in the branc hing pro cess b y a 10 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG factor of ( N − X 1 ( t )) / N . That is, if T ( t ) = Z t 0 N − X 1 ( s ) N ds ≤ t, then the t wo pro cesses can b e coupled s o that X 1 ( t ) = Y ( T ( t )), for all t ≥ 0. The time c hange will ha v e little effect as long as X 1 ( t ) is o ( N ) . T h e n ext lemma shows that on the r elev ant time scale, the n u m b er of mutan ts sta ys small with h igh probabilit y . Lemma 2.1. Fix t > 0 , ε > 0 , and let M t = max 0 ≤ s ≤ t/ ( N u 1 √ u 2 ) X 1 ( s ) . We have lim N →∞ P ( M t > εN ) = 0 . Pr oof. Since mutan t individuals giv e birth and die at th e same rate, the pro cess { X 1 ( s ) , s ≥ 0 } is a sub martingale. Because the rate of t yp e 1 m u tations is alw a ys b oun ded ab o ve by N u 1 , we ha v e E X 1 ( s ) ≤ N u 1 s f or all s . By Doob’s maximal inequalit y , P ( M t > εN ) ≤ E X 1 ( t/ N u 1 √ u 2 ) εN ≤ t εN √ u 2 , whic h go es to zero as N → ∞ , since N √ u 2 → ∞ . Using the time c hange in ( 2.1 ), w e ha v e P ( τ 2 > t/ N u 1 √ u 2 ) = E exp − u 2 Z t/ N u 1 √ u 2 0 Y ( T ( s )) ds . Changing v ariables r = T ( s ), wh ic h means s = U ( r ), where U = T − 1 , ds = U ′ ( r ) dr and the abov e is P ( τ 2 > t/ N u 1 √ u 2 ) = E exp − u 2 Z T ( t/ N u 1 √ u 2 ) 0 Y ( r ) U ′ ( r ) dr . When M t ≤ N ε , 1 ≥ T ′ ( t ) ≥ 1 − ε , so the inv erse function h as slop e 1 ≤ U ′ ( r ) ≤ 1 / (1 − ε ). Th us, in view of Lemma 2.1 , it is enough to prov e the result for the br anc hin g p ro cess, Y ( t ). Use Q to denote the distribution of { Y ( t ) , t ≥ 0 } , and let Q 1 denote the la w of th e pro cess starting from a single t yp e 1 and mo dified to hav e n o further m utations to t yp e 1. W e first compu te g 2 ( t ) = Q 1 ( τ 2 ≤ t ). W o darz and Komaro v a [ 24 ] do this, see pages 37–39, by using K olmogoro v’s forward equation to get a partial differen tial equation ∂ φ ∂ t ( t, y ) = ( y 2 − (2 + u 2 ) y + 1) ∂ φ ∂ y ( t, y ) W AITIN G TIMES F OR MUT A TIONS 11 for the generating function φ ( t, y ) = P j Q 1 ( X 1 ( t ) = j, X 2 ( t ) = 0) y j of the system in which t yp e 2’s are not allo w ed to giv e birth or d ie. They use the metho d of c haracteristics to reduce the PDE to a Riccati ordinary d ifferential equation. T o help readers who w an t to follo w th eir deriv ation, w e note that the last equation on page 38 is missing a factor of j in the last term and in the c hange of v ariables from y to z on page 39, 2 should b e r . Here, w e will u se Kolmogoro v’s b ac kwa rd different ial equation to deriv e an ODE, wh ic h has the adv ant age that it generalizes easily to the m stage problem. By considering what happ ens b et we en time 0 and h , g 2 ( t + h ) = g 2 ( t )[1 − (2 + u 2 ) h ] + h [2 g 2 ( t ) − g 2 ( t ) 2 ] + h · 0 + u 2 h · 1 + o ( h ) , where the four terms corresp ond to nothing happ ening, a birth, a death and a mutatio n of the original t yp e 1 to t yp e 2. Doing s ome algebra and lett ing h → 0 g ′ 2 ( t ) = − u 2 g 2 ( t ) − g 2 ( t ) 2 + u 2 . (2.2) If w e let r 1 > r 2 b e the solutions of x 2 + u 2 x − u 2 = 0 , that is, r i = − u 2 ± q u 2 2 + 4 u 2 2 , (2.3) w e can write this as g ′ 2 ( t ) = − ( g 2 ( t ) − r 1 )( g 2 ( t ) − r 2 ) . No w g 2 ( ∞ ) b e th e p r obabilit y that a type 2 offspring is ev entuall y generated in th e branching pro cess. Letting t → ∞ in ( 2.2 ) and noticing that t 7→ g 2 ( t ) is increasing implies g ′ 2 ( t ) → 0 , we s ee that 0 = − u 2 g 2 ( ∞ ) − g 2 ( ∞ ) 2 + u 2 , so 0 ≤ g 2 ( t ) < r 1 for all t and we h a ve 1 = g ′ 2 ( t ) ( r 1 − g 2 ( t ))( g 2 ( t ) − r 2 ) = 1 r 1 − r 2 g ′ 2 ( t ) g 2 ( t ) − r 2 + g ′ 2 ( t ) r 1 − g 2 ( t ) . In tegrating ln( g 2 ( t ) − r 2 ) − ln ( r 1 − g 2 ( t )) = ( r 1 − r 2 ) t − ln A, where A is a constan t that will b e c hosen later, so we h a ve g 2 ( t ) − r 2 r 1 − g 2 ( t ) = (1 / A ) e ( r 1 − r 2 ) t . A little alg ebra giv es g 2 ( t ) = r 1 + Ar 2 e ( r 2 − r 1 ) t 1 + Ae ( r 2 − r 1 ) t . 12 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG W e h a ve g 2 (0) = 0 , so A = − r 1 /r 2 and g 2 ( t ) = r 1 (1 − e ( r 2 − r 1 ) t ) 1 − ( r 1 /r 2 ) e ( r 2 − r 1 ) t . T o prepare for the asymp totics n ote th at ( 2.3 ) and the assumption that u 2 → 0 imply that r 1 − r 2 = q u 2 2 + 4 u 2 ∼ 2 √ u 2 , r 1 ∼ √ u 2 and r 1 /r 2 → − 1 so g 2 ( t ) ≈ √ u 2 (1 − e − 2 √ u 2 t ) 1 + e − 2 √ u 2 t or to b e p recise, if t √ u 2 → s, then g 2 ( t ) ∼ √ u 2 · 1 − e − 2 s 1 + e − 2 s . (2.4) Lemma 2.2. The waiting time for the first typ e 2 in the br anching pr o- c ess with immigr ation when e ach typ e 1 individual exp erienc es mutations at r ate N u 1 satisfies Q ( τ 2 ≤ t ) = 1 − exp − N u 1 Z t 0 Q 1 ( τ 2 ≤ s ) ds . (2.5) Pr oof. T yp e 1 m u tations are a Po isson pro cess with rate N u 1 . A p oint at time t − s is a success, that is, pro duces a type 2 b efore time t with probabilit y Q 1 ( τ 2 ≤ s ). By results for thinn in g a Poisson pr o cess, the n u m b er of successes by time t is Poisson with mean N u 1 R t 0 Q 1 ( τ 2 ≤ s ) ds . The result follo ws from the observ ation that Q ( τ 2 ≤ t ) is th e probabilit y of at least one success in the Poisson pro cess. T o find th e densit y fu nction, w e recall g 2 ( t ) = Q 1 ( τ 2 ≤ t ) and differen tiate to get N u 1 g 2 ( t ) exp − N u 1 Z t 0 g 2 ( s ) ds . Changing v ariables the density fu nction f 2 of τ 2 · N u 1 √ u 2 is giv en b y f 2 ( t ) = g 2 ( t/ N u 1 √ u 2 ) √ u 2 exp − N u 1 Z t/ N u 1 √ u 2 0 g 2 ( s ) ds . Changing v ariables r = sN u 1 √ u 2 in the integ r al the ab o v e is f 2 ( t ) = g 2 ( t/ N u 1 √ u 2 ) √ u 2 exp − Z t 0 g 2 ( r / N u 1 √ u 2 ) √ u 2 dr . W AITIN G TIMES F OR MUT A TIONS 13 If N u 1 → 0 , then ( 2.4 ) implies that the ab o ve con verges to exp( − t ) . If N u 1 → λ, the limit is h ( t ) exp( − R t 0 h ( s ) ds ) wh ere h ( s ) = 1 − e − 2 s/λ 1 + e − 2 s/λ , whic h completes the pro of of Theorem 1 . 3. A t wo -t yp e mo del. W e collect here some results for a simple t w o-t yp e p opulation mo del, whic h w e call mod el M 0 . W e assume that all individu als are either t yp e 0 or t yp e 1, and the p opulation size is alw a ys N . There are no m u tations, and the p opulation ev olve s acc ording to the Moran mo d el, so eac h in dividual dies at rate 1 and then is r ep laced b y a rand omly c hosen individual in the p opulation. Usually w e will assu me that the pro cess starts with jus t one t yp e 1 individual at time zero, but o ccasionally w e will also need to consider starting the pro cess w ith j typ e 1 ind ividuals. Denote by P j and E j probabilities and exp ectations when the pro cess is started with j t yp e 1 ind ivid uals, and write P = P 1 and E = E 1 . Let X ( t ) d enote the n u m b er of typ e 1 ind ividuals at time t . Let T k = inf { t : X ( t ) = k } b e the first time at whic h there are k t yp e 1 individuals, and let T = m in { T 0 , T N } b e the first time at whic h all individ- uals ha ve the s ame t yp e. Let L k b e the amoun t of time for whic h there are k t yp e one ind ividuals, which is the L eb esgue measure of { t < T : X ( t ) = k } . Let R k b e the num b er of times that the num b er of t yp e 1 individuals jumps to k from k − 1 or k + 1. Let R = 1 + P N − 1 k =1 R k b e the total num b er of births and deaths of t yp e 1 individuals. Du rrett and Schmidt [ 8 ] studied this mo del and sho wed th at E [ R k | T 0 < T N ] = 2( N − k ) 2 N ( N − 1) (3.1) and E [ R k | T N < T 0 ] = 2 k ( N − k ) N . (3.2) Equation ( 3.1 ) is (16) of [ 8 ], w h ile ( 3.2 ) comes from the b eginn in g of the pro of of L emm a 3 in [ 8 ]. Because P ( T N < T 0 ) = 1 / N , it follo ws from ( 3.1 ) and ( 3.2 ) that E [ R k ] = ( N − 1) E [ R k | T 0 < T N ] + E [ R k | T N < T 0 ] N = 2( N − k ) N ≤ 2 (3.3) and, therefore, E [ R ] = 1 + N − 1 X k =1 E [ R k ] ≤ 2 N . (3.4) 14 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG If 1 ≤ j ≤ N − 1, then letting A den ote the ev en t that there are at least j t yp e 1 individuals at some time, ( 3.4 ) giv es E j [ R ] = E j [ R 1 A ] ≤ E [ R 1 A ] P ( A ) = j E [ R 1 A ] ≤ j E [ R ] ≤ 2 j N . (3.5) T urning to the qu an tities L k , note that when there are k t yp e 1 ind ividuals, births and deaths are eac h happ ening at rate k ( N − k ) / N , so th e n umb er of t yp e 1 individu als c h anges again after an exp onen tial time with mean N/ [2 k ( N − k )]. Therefore, ( 3.3 ) giv es E [ L k ] = N 2 k ( N − k ) E [ R k ] = 1 k . (3.6 ) Since P j ( T k < T 0 ) = j /k for 1 ≤ j < N , w e h av e E j [ L k ] ≤ E 1 [ L k | T k < T 0 ] = E 1 [ L k ] P 1 ( T k < T 0 ) = 1 , (3.7) where to emphasize the change in initial cond ition, we ha v e wr itten E as E 1 . S ince T = P N − 1 k =1 L k , it also follo ws f rom ( 3.6 ) that E [ T ] = N − 1 X k =1 1 k ≤ C log N (3.8) and it follo ws from ( 3.7 ) that for j = 1 , . . . , N − 1, E j [ T ] ≤ N . (3.9) Finally , w e will use b ranc h ing pro cess theory to obtain the follo wing com- plemen t to ( 3.8 ). Lemma 3.1. Ther e exists a c onstant C such that P ( T > t ) ≤ C /t for al l 0 ≤ t ≤ N . Pr oof. Consider a cont in u ous-time branching pro cess s tarted with one individual in whic h eac h individual dies at rate one and giv es birth at rate one. Let T ′ b e the time at whic h the p ro cess b ecomes extinct. By a th eorem of Kolmogoro v [ 16 ], pr o ved in Section I .9 of [ 3 ], and the fact that a Mark ovia n con tinuous-time branc hing p ro cess can b e r educed to a discrete time Galton– W atson p ro cess b y only examining it at in teger times, we see that there is a constan t C ′ suc h that P ( T ′ > t ) ≤ C ′ /t for all t ≥ 0 . When th er e are k individuals in the branc hing p ro cess, b irths and deaths happ en at rate k . When ther e are k individuals in the mo del M 0 , b irths and d eaths happ en at r ate k ( N − k ) / N , whic h is at least k / 2 as long as k ≤ N/ 2. Sin ce the probabilit y that the n u m b er of individ uals in mo del M 0 ev er excee ds N/ 2 is at most 2 / N , w e h a ve P ( T > t ) ≤ 2 C ′ /t + 2 / N for all t , whic h implies the result. W AITIN G TIMES F OR MUT A TIONS 15 4. The p robabilit y of a t y p e m descendan t. W e no w consider mod el M 1 , whic h ev olv es in the same w a y as the pro cess describ ed in the In tro duction except that initial ly there is one typ e 1 ind ividual and N − 1 t y p e 0 indi- viduals, and n o fu rther t yp e 1 m utations occur . The n um b er of individu als of nonzero type in mo d el M 1 therefore ev olv es exactly lik e the n u mb er of t yp e 1 ind ividuals in mo del M 0 , defined at the b eginning of the previous section, but in mo del M 1 m u tations to t yp es greater than one are p ossible. The pr obabilit y , whic h w e denote by q m , that a type m individual is ev en- tually b orn in mo d el M 1 is the same as th e probabilit y that a giv en type one individu al in the pro cess describ ed in the In tro duction has a t y p e m descendan t. Our main goal in this sectio n is to pro ve the follo wing result. Pr oposition 4.1. Fix an inte ger m ≥ 2 . A ssume c onditions (ii), (iii) and (iv) of The or em 2 hold. Then q m ∼ r 1 ,m . W e will use Prop osition 4.1 to prov e Theorem 2 . T o prov e Theorem 3 , w e will need the follo wing corollary . Here w e denote by q j,m the p robabilit y that a t yp e m individual even tually app ears in a pro cess with initially one t yp e j ind ivid ual, N − 1 typ e 0 individuals, and m utations to t yp e 1 are not allo w ed . Corollar y 4.1. Fix an inte ger m ≥ 2 . Assume c onditions (ii) and (iii) of The or em 2 hold and that ( N r 1 ,m ) 2 → γ > 0 . Then q 2 ,m ∼ r 2 ,m . Pr oof. W e apply the m − 1 case of Prop osition 4.1 , w ith u 3 , . . . , u m in place of u 2 , . . . , u m − 1 . S in ce we are assuming (ii) and (iii), we need only to sho w th at N r 2 ,m → ∞ . Ho w ever, (ii) and (iii) imp ly N r 2 ,m N r 1 ,m = u 1 / 2 3 u 1 / 4 4 · · · u 1 / 2 m − 2 m u 1 / 2 2 u 1 / 4 3 · · · u 1 / 2 m − 2 m − 1 u 1 / 2 m − 1 m > b 1 / 2 2 b 1 / 4 3 · · · b 1 / 2 m − 2 m − 1 u − 1 / 2 m − 1 m → ∞ . This result and the assum p tion ( N r 1 ,m ) 2 → γ > 0 imply N r 2 ,m → ∞ . W e will prov e Prop osition 4.1 using a branching pro cess appro ximation. W e w ill approxima te mo del M 1 b y a cont in u ous-time m u lti-t yp e branc hing pro cess in whic h individ uals of type 1 ≤ j < m d ie at rate 1, give birth at rate 1 and m utate to individuals of t yp e j + 1 at rate u j +1 . L et p j,m b e the probabilit y that a type j individual ev entually has a descendan t of t yp e m in the branc hing pro cess and let p m = p 1 ,m . Lemma 4.1. If c onditions (ii) and (iii) of The or em 2 hold, then p j,m ∼ r j,m . 16 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG Pr oof. W e pro ceed b y induction starting at j = m and working do w n to j = 1. C learly , p m,m = 1 , so th e result is v alid f or j = m . Now assu me the result is true f or j + 1. By cond itioning on the first eve n t in the br an ching pro cess, it follo ws that p j,m = 1 2 + u j +1 (2 p j,m − p 2 j,m ) + u j +1 2 + u j +1 p j +1 ,m . Multiplying by 2 + u j +1 and rearranging, we get p 2 j,m + bp j,m − u j +1 p j +1 ,m = 0, where b = u j +1 . The only p ositiv e solution is p j,m = − b + q b 2 + 4 u j +1 p j +1 ,m 2 . (4.1) Calculus tells that f or h > 0 √ x + h − √ x = Z x + h x 1 2 √ y dy ≤ h 2 √ x , so, w e ha v e 2 √ u j +1 p j +1 ,m ≤ q 4 u j +1 p j +1 ,m + b 2 (4.2) ≤ 2 √ u j +1 p j +1 ,m + b 2 4 √ u j +1 p j +1 ,m . Conditions (ii) and (iii) imp ly that u j +1 ≪ r j +1 ,m and, therefore, that √ u j +1 r j +1 ,m ≫ b = u j +1 . Since p j +1 ,m ∼ r j +1 ,m b y the ind uction hypoth- esis, it follo ws f r om ( 4.1 ) and ( 4.2 ) that p j,m ∼ √ u j +1 r j +1 ,m . T he lemma follo ws by ind uction. Remark. One gets the same result for a n u m b er of other v ariants of the mo del. W e lea v e it to the reader to c hec k th at Lemma 4.1 holds w hen m u tation only o ccurs at b irth. T o prepare for the p ro of of Lemma 4.7 , w e will no w sho w that it holds when t yp e j ’s giv e birth to t yp e j ’s at rate one and to typ e j + 1 ’s at rate u j +1 . I n this case, the first equ ation is p j,m = 1 2 + u j +1 (2 p j,m − p 2 j,m ) + u j +1 2 + u j +1 ( p j,m + p j +1 ,m − p j,m p j +1 ,m ) and rearranges to b ecome p 2 j,m + u j +1 p j +1 ,m p j,m − u j +1 p j +1 ,m = 0. T aking b = u j +1 p j +1 ,m , th e pro of goes as b efore. W e w ill no w pro ve Prop osition 4.1 b y ind uction. W e b egin with the case m = 2 , in whic h the comparison with the branching pro cess is straigh tfor- w ard . W AITIN G TIMES F OR MUT A TIONS 17 Lemma 4.2. Under the assumptions of Pr op osition 4.1 with m = 2 , we have q 2 ∼ r 1 , 2 = u 1 / 2 2 . Pr oof. If we trac k the num b er of t yp e 1 in dividuals in mo del M 1 b efore the firs t t yp e 2 mutatio n occurs, up w ard and do wnw ard jumps o ccur at the same rate, whic h if there are k type 1 individuals is k ( N − k ) / N . F or the branc hing pro cess, wh en there are k t yp e 1 in dividuals, upw ard and do w n- w ard jumps occur at rate k . Th erefore, the em b edd ed jump c hain (which giv es the sequence of states visited by the contin uous-time c hain) is a sim- ple r andom w alk S n with S 0 = 1 b oth for mod el M 1 and for the branc hing pro cess. Th erefore, writing p 2 as a function of th e underlying m u tation rate, w e claim that for an y L , p 2 ( u 2 ) − 1 / N ≤ q 2 ≤ p 2 ( u 2 N/ ( N − L )) + 1 /L. (4.3) The first inequalit y follo ws from the fact that unless th e num b er of t yp e 1 individuals in mo del M 1 reac hes N , whic h h app ens with probabilit y 1 / N , mo del M 1 has the same embedd ed jump c h ain as the branc h ing pro cess and jumps more slo wly . F or the second inequalit y , w e n ote that the pr ob ab ility the Moran mo del reac h es heigh t L is 1 /L . When this do es not o ccur , the Moran mo d el alw ays jump s at rate at least ( N − L ) / N times the branching pro cess r ate. Lemma 4.1 giv es p 2 ( u 2 ) ∼ u 1 / 2 2 . Condition (iv) gives N u 1 / 2 2 → ∞ , so w e can choose L suc h that L / N → 0 and Lu 1 / 2 2 → ∞ . Under these conditions, ( 4.3 ) implies q 2 ∼ u 1 / 2 2 . F or the r est of this section, w e will fix m and assume that the assumptions of Prop osition 4.1 hold. W e will also assume th at Prop osition 4.1 has b een established for m − 1, wh ic h imp lies that q 2 ,m ∼ r 2 ,m . W e will reduce the general case to the m = 2 case in whic h t yp e 2 mutati ons o ccur at rate u 2 r 2 ,m . The next t w o lemmas w ill allo w us to ignore certa in t yp e 2 mutati ons. Lemma 4.3. L et A m b e the event that in mo del M 1 some typ e 2 mutation that o c curs while ther e is ano ther individual in the p opulation of typ e 2 or higher has a typ e m desc e ndant. Then P ( A m ) ≪ r 1 ,m . Pr oof. Let ε > 0. Let B b e the ev ent that the num b er of individu als in the p opu lation of t yp e 1 or higher n ev er exceeds ε − 1 r − 1 1 ,m , so P ( B c ) ≤ εr 1 ,m . Let U = { t : there is an individual of type 2 or higher alive at time t } . On B , t yp e 2 m u tations o ccur at rate at most ε − 1 r − 1 1 ,m u 2 and ha v e a t yp e m de- scendan t with probabilit y q 2 ,m . Therefore, letting | U | denote the L eb esgue measure of U , w e hav e P ( A m ) ≤ εr 1 ,m + E [ | U | 1 B ] ε − 1 r − 1 1 ,m u 2 q 2 ,m . 18 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG F or k ≤ ε − 1 r − 1 1 ,m , it follo ws fr om ( 3.6 ) that th e exp ected amoun t of time for whic h there are k ind ividuals of type 1 or higher is 1 /k , and so the exp ected n u m b er of type 2 mutat ions du ring this time is at most (1 /k )( k u 2 ) = u 2 . Therefore, the exp ected num b er of t yp e 2 m u tations while th ere are at most ε − 1 r − 1 1 ,m individuals of t yp e 1 or h igher is at most ε − 1 r − 1 1 ,m u 2 . By ( 3.8 ), the exp ected amoun t of time for whic h these mutatio ns or their off- spring are alive in the p opulation is at most ( C log N ) ε − 1 r − 1 1 ,m u 2 . Therefore, E [ | U | 1 B ] ≤ ( C log N ) ε − 1 r − 1 1 ,m u 2 . Since q 2 ,m ∼ r 2 ,m b y th e induction hyp oth- esis and u 2 r 2 ,m = r 2 1 ,m , it follo ws that there exists a constan t C such that P ( A m ) ≤ εr 1 ,m + C (log N ) ε − 2 r − 2 1 ,m u 2 2 r 2 ,m = εr 1 ,m + C (log N ) ε − 2 u 2 . Conditions (ii) an d (iii) imp ly that there exist constan ts C 1 and C 2 suc h that (log N ) u 2 r 1 ,m ≤ C 1 u 1 / 2 m − 1 2 log N ≤ C 2 u 1 / 2 m − 1 m log N → 0 . It follo ws that lim sup N →∞ r − 1 1 ,m P ( A m ) ≤ ε, whic h implies the lemma. Lemma 4.4. L et ε > 0 . L et B m b e the event that i n mo del M 1 some typ e 2 mutation that o c curs while ther e ar e fewer than εr − 1 1 ,m individuals in the p opulation of typ e 1 or higher has a typ e m desc endant. Then ther e is a c onstant C , not dep ending on ε , such that P ( B m ) ≤ C εr 1 ,m . Pr oof. As n oted in the pro of of Lemma 4.3 , the expected n um b er of t yp e 2 mutati ons while there are k individuals of typ e 1 or higher is u 2 . Therefore, the exp ected num b er of t yp e 2 m utations while there are few er than εr − 1 1 ,m individuals of t yp e 1 or h igher is at most εr − 1 1 ,m u 2 . By th e in- duction hyp othesis, eac h su c h m u tation pro du ces a type m d escendan t with probabilit y q m ∼ r 2 ,m , so the probabilit y that one of these m utations pro- duces a t yp e 2 descendan t is at most C εr − 1 1 ,m r 2 ,m u 2 . T he desired result no w follo ws f r om the fact that u 2 r 2 ,m = r 2 1 ,m . Our strategy is to show that we can reduce the p roblem to the m = 2 case b y assuming that eac h t yp e 2 mutati on in d ep end ently generates a type m W AITIN G TIMES F OR MUT A TIONS 19 descendan t w ith probabilit y q 2 ,m . Complicating this pictur e is the fact that the evo lution of the num b er of type 1 ind ividuals (which pro duce the t yp e 2 m u tations) is not indep endent of the success of the t yp e 2 mutatio ns b ecause a new individ ual of t yp e j ≥ 2 ma y replace an existing t yp e 1 individu al and vice v ersa. T o sho w that this is not a s ignifi can t p roblem, we w ill construct a coupling of mo del M 1 with another pr o cess in wh ic h this d ep end ence has b een eliminated. W e first define mo del M 2 to evolv e lik e mo del M 1 except that initially there are k individ uals of t yp e 1 and N − k of typ e 0, and t yp e 2 muta tions are only p ermitted when there are no ind ividuals of typ e j ≥ 2. W e th en compare mo del M 2 to mo del N 2 , in whic h the type 1 individu als are decoupled from t yp e 2 individuals and their offspring b y d eclaring that (pro vid ed a typ e 0 individu al exists): • if a prop osed mo v e exc hanges a type 1 and a type j ≥ 2, we in stead exc hange a t y p e 0 and a t yp e j ; • a m utation that o ccurs to a t yp e 1 pr o duces a n ew type 2 individual but replaces a typ e 0 ind ividual instead of the t yp e 1 that mutat ed. T o define the couplin g precisely , in tro duce a Poisson pro cess with r ate N at w h ic h the successiv e exc hanges will o ccur and let i n and j n b e ind ep endent i.i.d. un iform on { 1 , 2 , . . . , N } . I n b oth mo dels, we replace individual i n with a cop y of individu al j n . In mo del N 2 , if i n has t yp e 1 and j n has t yp e 2 or higher, then we c ho ose a t yp e 0 ind ividual at rand om to b ecome typ e 1, so that the num b er of type 1 ind ividuals sta ys the same. Like wise, if i n has t yp e 2 or higher and j n has t yp e 1, then w e c ho ose a type 1 in d ividual to b ecome t yp e 0 in mo del N 2 . This recip e breaks do wn when there are n o individuals of t yp e 0. Ho wev er, Lemma 4.5 sh ows th at with h igh probability the num b er of individuals of n on zero typ e is o ( N ) up to time τ m . F or the m u tations, we ha v e for eac h 1 ≤ i ≤ N a P oisson pro cess w ith r ate u 2 , whic h in b oth mo dels causes a mutatio n of the i th individu al, unless either the i th individual h as typ e 0 or the i th individual h as type 1 and there is an individual of t yp e 2 or higher in the p opulation. In mo del N 2 , if a t yp e 1 individual mutate s to type 2, a type 0 individ u al is c hosen at r andom to b ecome t yp e 1, to ke ep the num b er of t yp e 1 individuals constan t. Let X 1 ( t ) and Y 1 ( t ) b e the n um b er of type 1 individ uals at time t in mo dels M 2 and N 2 , resp ectiv ely . Let Z ( t ) = X 1 ( t ) − Y 1 ( t ). Let ˆ X 2 ( t ) and ˆ Y 2 ( t ) b e the num b er of individ u als in mo d els M 2 and N 2 , resp ectiv ely , of t yp e greate r than or equal to 2. Note that by renum b erin g the ind ivid uals as the pr o cess ev olves if necessary , w e can ensur e that for all t ≥ 0, at time t there are min { X 1 ( t ) , Y 1 ( t ) } in tegers j suc h that the j th individual has t yp e 1 in b oth mo del M 2 and mo del N 2 . Note also that with the ab o v e couplin g, if a t yp e 2 mutatio n o ccurs at the same time in b oth mo dels, descendant s of this mutat ion will al w ays ha ve the same type in b oth mo dels. This means 20 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG that if the mutatio n has a t yp e m descendant in one m o del, then it will ha ve a t yp e m descendan t in the other. Finally , as long as the n umb er of individuals of nonzero t yp e sta ys b elo w N / 2, we can also ensu r e that there is no j suc h that the j th individual h as typ e 1 in one of the t wo mo d els and t yp e 2 or higher in the other. The lemma b elo w, combined with condition (iv), ensu res that in b oth mo dels, the num b er of in dividuals of n onzero type sta ys muc h smaller than N . Lemma 4.5. Fix t > 0 . Supp ose X 1 (0) = Y 1 (0) = [ εr − 1 1 ,m ] and ˆ X 2 (0) = ˆ Y 2 (0) = 0 . Assume f is a fu nction of N such that f ( N ) r 1 ,m → ∞ as N → ∞ . Then using → p to denote c onver ge nc e in pr ob ability, we have max 0 ≤ s ≤ tr − 1 1 ,m X 1 ( s ) + ˆ X 2 ( s ) f ( N ) → p 0 and max 0 ≤ s ≤ tr − 1 1 ,m Y 1 ( s ) + ˆ Y 2 ( s ) f ( N ) → p 0 . Pr oof. In mo d el M 2 , individuals of type 1 or higher giv e birth and d ie at the same rate, so ( X 1 ( s ) + ˆ X 2 ( s ) , s ≥ 0) is a martingale and E [ X 1 ( tr − 1 1 ,m ) + ˆ X 2 ( tr − 1 1 ,m )] = X 1 (0) + ˆ X 2 (0) = [ εr − 1 1 ,m ] . By Doob’s maximal inequalit y , if δ > 0 , th en P max 0 ≤ s ≤ tr − 1 1 ,m X 1 ( s ) + ˆ X 2 ( s ) f ( N ) > δ ≤ E [ X 1 ( tr − 1 1 ,m ) + ˆ X 2 ( tr − 1 1 ,m )] δ f ( N ) ≤ εr − 1 1 ,m δ f ( N ) → 0 as N → ∞ , whic h implies the fi rst statemen t of the lemma. In mo del N 2 , muta tions of t yp e 1 individu als cause new t yp e 2 individuals to replace t yp e 0 in dividuals. Births and deaths o ccur at the same rate, so the pro cess ( Y 1 ( s ) , s ≥ 0) is a martingale, while ( Y 1 ( s ) + ˆ Y 2 ( s ) , s ≥ 0) is a submartingale. Now E [ Y 1 ( s )] = [ εr − 1 1 ,m ] for all s , so the exp ected num b er of t yp e 2 ind ividuals that app ear b efore time tr − 1 1 ,m b ecause of mutation is at most εr 1 ,m · tr − 1 1 ,m · u 2 = εu 2 r − 2 1 ,m t . It f ollo ws that E [ Y 1 ( tr − 1 1 ,m ) + ˆ Y 2 ( tr − 1 1 ,m )] ≤ εr − 1 1 ,m + εu 2 r − 2 1 ,m t. No w u 2 r − 1 1 ,m = u 2 u 1 / 2 2 u 1 / 4 2 · · · u 1 / 2 m − 1 m = u 1 − 1 / 2 m − 1 2 u 1 / 2 2 u 1 / 4 2 · · · u 1 / 2 m − 1 m · u 1 / 2 m − 1 2 → 0 , (4.4) W AITIN G TIMES F OR MUT A TIONS 21 b ecause condition (ii) implies that the first factor is b ounded by a constan t, so Doob’s maximal inequalit y this time giv es P max 0 ≤ s ≤ tr − 1 1 ,m Y 1 ( s ) + ˆ Y 2 ( s ) f ( N ) > δ ≤ εr − 1 1 ,m + εu 2 r − 2 1 ,m t δ f ( N ) → 0 , whic h implies the second half of the lemma. W e n o w work on b ound ing the pro cess ( Z ( t ) , t ≥ 0). There are three typ es of even ts th at cause this pr o cess to jump. First, whenever a t yp e 1 individual in mo del M 2 m u tates to t yp e 2, there is no corresp onding c hange in mo d el N 2 , b ecause an y new type 2 ind ividual in mo del N 2 resulting from mutat ion replaces a t yp e 0. These c hanges cause the Z pro cess to decrease b y one. Letting µ ( t ) b e the rate at w hic h they are occurr ing at time t , w e ha v e 0 ≤ µ ( t ) ≤ u 2 X 1 ( t ) , where the second inequalit y could b e strict b ecause m utations are supp ressed if there is already a t yp e 2 in d ividual in the p op u lation. Second, one of the “extra” | Z ( t ) | t yp e 1 individuals in one pro cess or the other could exp erience a birth or a death. This could cause the Z pro cess to increase or decrease by one. If X 1 ( t ) > Y 1 ( t ), then at time t , b oth in- creases and decreases in th e Z p ro cess o ccur b ecause of suc h changes at rate | Z ( t ) | ( N − | Z ( t ) | ) / N , b ecause the Z pro cess change s u nless the other indi- vidual in volv ed in the exc hange wa s also one of the | Z ( t ) | ind ividuals that are type 1 in mo del M 2 but not mod el N 2 . If Y 1 ( t ) > X 1 ( t ), then increases and decreases in th e Z pro cess occur at r ate | Z ( t ) | ( N − | Z ( t ) | − ˆ Y 2 ( t )) / N b ecause exc hanges b etw een a t yp e 1 ind ivid ual and an individual of t yp e 2 or higher are prohib ited in mo d el N 2 . Finally , th ere are transitions in which one of the min { X 1 ( t ) , Y 1 ( t ) } in di- viduals that are t yp e 1 in b oth pro cesses exp eriences a b irth or death, but the other individual inv olv ed in the exc hange is one of the ˆ Y 2 ( t ) in d ividuals that has t yp e 2 in m o del N 2 , so the t yp e 1 p opulation do es not c hange in mo del N 2 . S uc h change s o ccur at rate ˆ Y 2 ( t ) min { X 1 ( t ) , Y 1 ( t ) } / N . Th us, if we let λ ( t ) = | Z ( t ) | ( N − | Z ( t ) | − ˆ Y 2 ( t ) 1 { Y 1 ( t ) >Z 1 ( t ) } ) N + ˆ Y 2 ( t ) min { X 1 ( t ) , Y 1 ( t ) } N , then at time t th e Z pro cess is in creasing b y 1 at r ate λ ( t ) and decreasing by 1 at rate λ ( t ) + µ ( t ) . The next result u ses these facts to con trol the difference b et w een X 1 ( t ) and Y 1 ( t ). Lemma 4.6. Fix t > 0 . L et Z N ( s ) = r 1 ,m Z ( sr − 1 1 ,m ) for al l s ≥ 0 . If X 1 (0) = Y 1 (0) = εr − 1 1 ,m and ˆ X 2 (0) = ˆ Y 2 (0) = 0 , then max 0 ≤ s ≤ t Z N ( s ) → p 0 . 22 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG Pr oof. W e w ill use Theorem 4.1 from Chapter 7 in [ 10 ] to sho w that Z N con verge s to a d iffusion with b ( x ) = 0, a ( x ) = 2 | x | , and initial p oin t 0, so the limit is identic ally zero. The first step is to observe that the Y amada– W atanab e theorem; see, for example, (3.3) on page 193 of [ 6 ], giv es p ath wise uniqueness for the limiting SDE, whic h in turn implies that the martingale problem is well p osed. T o verify the other assump tions of the theorem, define B N ( t ) = − Z t 0 µ ( sr − 1 1 ,m ) ds and A N ( t ) = Z t 0 r 1 ,m (2 λ ( sr − 1 1 ,m ) + µ ( s r − 1 1 ,m )) ds. In view of the transition rates f or the pr o cess ( Z ( t ) , t ≥ 0), we see that at time s the pro cess Z N ( s ) exp eriences p ositiv e jumps by th e amount r 1 ,m at rate λ ( sr − 1 1 ,m ) r − 1 1 ,m and negativ e jump s by the s ame amount at rate ( λ ( sr − 1 1 ,m ) + µ ( sr 1 ,m )) r − 1 1 ,m . Th erefore, letting M N ( t ) = Z N ( t ) − B N ( t ), the pro cesses ( M N ( t ) , t ≥ 0) and ( M 2 N ( t ) − A N ( t ) , t ≥ 0) are martingales. T o obtain the result of the lemma from Theorem 4.1 in Chapter 7 of [ 10 ], it remains to sho w that for any fixed T > 0, we ha v e sup 0 ≤ t ≤ T | B N ( t ) | → p 0 (4.5) and sup 0 ≤ t ≤ T A N ( t ) − Z t 0 2 | Z N ( s ) | ds → p 0 . (4.6) T o pro ve ( 4.5 ), note th at sup 0 ≤ t ≤ T | B N ( t ) | ≤ T sup 0 ≤ t ≤ T µ ( tr − 1 1 ,m ) ≤ T u 2 max 0 ≤ t ≤ T r − 1 1 ,m X 1 ( t ) . Since r 1 ,m / ( T u 2 ) → ∞ by ( 4.4 ), ( 4.5 ) no w follo w s from Lemma 4.5 with f ( N ) = 1 / ( T u 2 ). F or ( 4.6 ), note that A N ( t ) − Z t 0 2 | Z N ( s ) | ds = r 1 ,m Z t 0 − 2 | Z ( sr − 1 1 ,m ) | 2 N − 2 | Z ( sr − 1 1 ,m ) | ˆ Y 2 ( sr − 1 1 ,m ) 1 { Y 1 ( sr − 1 1 ,m ) >Z 1 ( sr − 1 1 ,m ) } N + 2 ˆ Y 2 ( sr − 1 1 ,m ) min { X 1 ( sr − 1 1 ,m ) , Y 1 ( sr − 1 1 ,m ) } N + µ ( sr − 1 1 ,m ) ds. It suffices to control the absolute v alues of the four terms o v er all t ≤ T . Note that Z ( s r − 1 1 ,m ) ≤ m ax { X 1 ( sr − 1 1 ,m ) , Y 1 ( sr − 1 1 ,m ) } . Therefore, b y Lemma 4.5 W AITIN G TIMES F OR MUT A TIONS 23 with f ( N ) = q N/r 1 ,m , the three quan tities max 0 ≤ s ≤ T r − 1 1 ,m r 1 / 2 1 ,m N − 1 / 2 | Z ( s ) | , max 0 ≤ s ≤ T r − 1 1 ,m r 1 / 2 1 ,m N − 1 / 2 ˆ Y 2 ( s ) and max 0 ≤ s ≤ T r − 1 1 ,m r 1 / 2 1 ,m N − 1 / 2 X 1 ( s ) all con v erge in pr obabilit y to zero as N → ∞ . This is enough to establish the con ve r gence of the firs t three terms. Th e result for th e third term follo w s from ( 4.5 ) and the fact th at r 1 ,m → 0. In the mod el N 2 , t yp es j ≥ 2 hav e the same relationship to type 1 indi- viduals as in the br anc hin g p ro cess. T hat is, t yp e 1’s give birth to type 2’s, but the fate of a t yp e 2 family do es not affect th e num b er of t yp e 1 indi- viduals b ecause a t yp e 1 individu al cannot b e exc han ged w ith an in dividual of t yp e 2 or higher. L emma 4.3 has sho wn that we can ignore t yp e 2 birth s that occur when another t yp e 2 is present, so su ccessiv e t yp e 2 births giv e indep en d en t c h ances of pr o ducing a t yp e m ind ividual. W e are no w close to our goal announced in the In tro duction of reducing the m -t yp e problem to th e 2-t yp e problem with ¯ u 2 = u 2 q 2 ,m , that is, to the simplified mod el in whic h at eac h type 2 mutatio n, we fl ip a coin with pr ob ab ility q 2 ,m of heads to see if it will generate a typ e m ind ividual. Let mo del N ′ 2 b e mo del N 2 mo dified so that if a t yp e 2 m utation o ccurs when ˆ Y 2 ( t ) > 0 , instead of su ppressing this ev ent en tirely , we flip a coin with probabilit y q 2 ,m of heads. W e then add a t yp e m individual to the p opulation if the coin is heads and otherwise mak e no c hange. Lemma 4.3 im p lies th at the differen ce b et w een the probabilit y of getting a type m individual in mo del N 2 and the probabilit y of getting a t yp e m individual in mo d el N ′ 2 tends to zero as N → ∞ . Ho wev er, it is easier to p ro ve the next result using mo del N ′ 2 b ecause in mo del N ′ 2 , eac h typ e 1 ind ividual is giving rise to individuals that w ill pro duce a t yp e m descendant at rate u 2 q 2 ,m , regardless of whether there are other individuals in the p opu lation of t yp e 2 or higher. Lemma 4.7. L et ε > 0 . Consider mo del N ′ 2 starting fr om [ εr − 1 1 ,m ] typ e 1 individuals at time zer o. L et h 1 N ,m,ε b e the pr ob ability that a typ e m individual is b orn at some time. Then lim N →∞ h 1 N ,m,ε = 1 − e − ε . Pr oof. Consider a mo d ified branching pro cess in wh ich t yp e j individ - uals giv e birth at rate one, d ie at r ate one, and giv e birth to type j + 1 individuals at r ate u j +1 . Let h 0 N ,m,ε b e the pr obabilit y that if the br anc hin g pro cess starts with [ εr 1 ,m ] individuals, a t yp e m in dividual is b orn at some time. Since differen t families are indep endent, Lemma 4.1 im p lies h 0 N ,m,ε = 1 − (1 − p m ) [ εr − 1 1 ,m ] → 1 − e − ε , where p m is the pr obabilit y that a type 1 individual has a t yp e m descendan t. 24 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG W e n o w compare this pro cess to mo del N ′ 2 . The num b er of type 1 individ- uals in mo del N ′ 2 jumps m ore slowly than the n umb er of t yp e 1 individuals in th e branc h ing p ro cess, bu t in b oth pro cesses type 1 in d ividuals giv e b irth to t yp e 2 ind ividuals at rate u 2 , and th en typ e 2 in dividuals and their de- scendan ts ev olve indep enden tly of the t yp e 1’s. Therefore, if the probabilit y p 2 ,m that a t yp e 2 individual in the branching pro cess pr o duces a t yp e m descendan t we re equal to q 2 ,m , then it w ould follo w that h 1 N ,m,ε ≥ h 0 N ,m,ε . Instead, w e only hav e p 2 ,m ∼ q 2 ,m b ecause p 2 ,m ∼ r 2 ,m b y th e remark after Lemma 4.1 an d q 2 ,m ∼ r 2 ,m b y the induction h yp othesis. It follo ws that h 1 N ,m,ε ≥ h 0 N ,m,ε (1 − o (1)) → 1 − e − ε . T o get a b ound in the opp osite direction, observe that w e can pic k K → ∞ so that L = K r − 1 1 ,m = o ( N ), and with probabilit y tend ing to one as N → ∞ , the num b er of typ e 1’s do es n ot reac h L . Th erefore, writing h 1 N ,m,ε and h 0 N ,m,ε as fun ctions of the rate at which t yp e 1 ind ividuals giv e b irth to t yp e 2 individuals, w e hav e h 1 N ,m,ε ( u 2 ) ≤ h 0 N ,m,ε ( u 2 N/ ( N − L ))(1 + o (1)) + o (1) → 1 − e − ε , whic h completes the pro of. Lemma 4.8. L et ε > 0 . Consider mo del M 2 starting fr om [ εr − 1 1 ,m ] typ e 1 individuals at time zer o. L et h N ,m,ε b e the pr ob ability that a typ e m individual is b orn at some time. Then lim N →∞ | h N ,m,ε − h 1 N ,m,ε | = 0 . Pr oof. Recall the coup lin g b et ween mo d el M 2 and mo del N 2 describ ed earlier in th is section. With this coupling, if a t yp e 2 mutatio n o ccur s at the same time in b oth p ro cesses, then it pro duces a t yp e m descendant in one pro cess if and only if it pro d uces a t yp e m d escendan t in the other. Consequent ly , it suffices to b oun d the probabilit y that some t yp e 2 m utation that app ears in one pro cess bu t not the other pro du ces a t yp e m descendan t. Lemma 4.3 b ounds th is probabilit y for mutat ions that o ccur in one mo del but get su ppressed in the other b ecause there are no individ uals of t yp e 2 or higher. It remains to consider th e m utations exp erienced b y the | Z ( t ) | indi- viduals that are type 1 in one p ro cess but not the other. Pic k s large enough so that the probabilit y N 2 or M 2 do es not die out by time s r − 1 1 ,m is < δ . Pic k η so that η s < δ 2 . By Lemma 4.6 , if N is large, we ha v e max t ≤ s | Z N ( t ) | < η with pr obabilit y > 1 − δ . The exp ected num b er of bir ths that o ccur in one pro cess b ut not in the other b efore time sr − 1 1 ,m when m ax t ≤ s | Z N ( t ) | < η is b ound ed by 2 η r − 1 1 ,m · sr − 1 1 ,m u 2 ≤ 2 δ 2 r − 2 1 ,m u 2 . W AITIN G TIMES F OR MUT A TIONS 25 Using Cheb yshev’s inequalit y , it follo ws th at with probabilit y > 1 − 4 δ the n u m b er of t yp e 2 mutan t births that o ccur in one pro cess b ut n ot the other is b ounded by δ r − 2 1 ,m u 2 = δ r − 1 2 ,m . When this o ccurs, the success p r obabilities differ b y at most δ b ecause eac h mutatio n has p robabilit y q 2 ,m ∼ r 2 ,m of pro du cing a typ e m descendan t. Since δ > 0 is arbitrary , th e desired results follo w. Pr oof of Proposition 4.1 . The p robabilit y th at the num b er of indi- viduals of t yp e greater than zero reac hes [ εr − 1 1 ,m ] is 1 / [ εr − 1 1 ,m ]. If, at the time T when the num b er of ind ividuals of typ e greater than zero reac hes [ εr − 1 1 ,m ], w e change the t yp e of all individuals whose t yp e is nonzero to t yp e 1, and if w e disregard t yp e 2 m utations that occur when there is another individual of t yp e j ≥ 2 , then the probabilit y of getting a t yp e m ind ividual after th is time b ecomes h N ,m,ε . Since these c hanges of the typ es can only redu ce the probabilit y of getting a t yp e m individual, w e ha ve q m ≥ 1 [ εr − 1 1 ,m ] h N ,m,ε . (4.7) Also, for a t yp e m in d ividual to app ear, either the type m individual m u st b e descended from a t y p e 1 individu al that is alive at time T , or else the t yp e m individual m ust b e descended from a t yp e 2 individual that existed b efore time T , so using Lemmas 4.3 and 4.4 , it follo ws that q m ≤ 1 [ εr − 1 1 ,m ] h N ,m,ε + C εr 1 ,m . (4.8) The result f ollo ws by letting ε → 0. 5. Pro of of Theorem 2 . In this section, w e complete the pro of of The- orem 2 . T he argument is based on the follo wing result on Poi sson appro xi- mation, whic h is part of Theorem 1 of [ 2 ]. Lemma 5.1. Supp ose ( A i ) i ∈I is a c ol le ction of events, wher e I is any index set. L et W = P i ∈I 1 A i b e the numb er of events that o c cur, and let λ = E [ W ] = P i ∈I P ( A i ) . Supp ose for e ach i ∈ I , we have i ∈ β i ⊂ I . L et F i = σ (( A j ) j ∈I \ β i ) . De fine b 1 = X i ∈I X j ∈ β i P ( A i ) P ( A j ) , b 2 = X i ∈I X i 6 = j ∈ β i P ( A i ∩ A j ) , b 3 = X i ∈I E [ | P ( A i |F i ) − P ( A i ) | ] . Then | P ( W = 0) − e − λ | ≤ b 1 + b 2 + b 3 . 26 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG W e will use the follo wing lemma to get th e second momen t estimate needed to b ound b 2 . When we apply th is r esult, the individuals b orn at times t 1 and t 2 will b oth b e t yp e 1. W e call the second one t yp e 2 to b e ab le to easily distinguish the descendants of the t wo ind ividuals. Lemma 5.2. Fix times t 1 < t 2 . Consider a p opulation of size N which evolves ac c or ding to the Mor an mo del in which al l individuals initial ly have typ e 0. Ther e ar e no mutations, e xc ept that one individual b e c omes typ e 1 at time t 1 , and one typ e 0 individual (if ther e is one) b e c omes typ e 2 at time t 2 . Fix a p ositive inte ger L ≤ N/ 2 . F or i = 1 , 2 , let Y i ( t ) b e the numb er of typ e i individuals at time t and let B i b e the e v ent that L ≤ max t ≥ 0 Y i ( t ) ≤ N / 2 . Then P ( B 1 ∩ B 2 ) ≤ 2 /L 2 . Pr oof. Because ( Y 1 ( t ) , t ≥ t 1 ) is a m artingale, it is clear th at P ( B 1 ) ≤ 1 /L . Let s 1 < s 2 < · · · < s J b e the ordered times, after time t 2 , at wh ic h th e Y 1 pro cess jumps. F or t ≥ t 2 , let Z ( t ) = Y 2 ( t ) A ( t ), where A ( t ) = N − Y 1 ( t 2 ) N − Y 1 ( t ) = Y i : s i ≤ t N − Y 1 ( s i − ) N − Y 1 ( s i ) . W e claim that conditional on ( Y 1 ( t ) , t ≥ t 1 ), the pro cess ( Z ( t ) , t ≥ t 2 ) is a martingale. T o see th is, note that b etw een the times s i , b irths and d eaths of type 2 individuals occur at the same r ate, ev en conditional on ( Y 1 ( t ) , t ≥ t 1 ), so Z ( t ) exp eriences b oth p ositiv e and n egativ e ju mps of size ( N − Y 1 ( t 2 )) / ( N − Y 1 ( t )) at the same rate. A t the time s i , if Y 1 ( s i ) = Y 1 ( s i − ) + 1, then one of the N − Y 1 ( s i − ) individu als of t yp e other than 1 dies at time s i , so we ha ve Y 2 ( s i ) = Y 2 ( s i − ) − 1 with probabilit y α i = Y 2 ( s i − ) / ( N − Y 1 ( s i − )) and Y 2 ( s i ) = Y 2 ( s i − ) with probabilit y 1 − α i . Note that (1 − α i ) Y 2 ( s i − ) + α i ( Y 2 ( s i − ) − 1) = Y 2 ( s i − ) − α i = Y 2 ( s i − ) 1 − 1 N − Y 1 ( s i − ) = Y 2 ( s i − ) N − Y 1 ( s i ) N − Y 1 ( s i − ) . Lik ewise, if Y 1 ( s i ) = Y 1 ( s i − ) − 1 , then one of the N − Y 1 ( s i − ) individuals of t yp e other than 1 giv es bir th at time s i , s o Y 2 ( s i ) = Y 2 ( s i − ) + 1 with prob- abilit y α i = Y 2 ( s i − ) / ( N − Y 1 ( s i − )) and Y 2 ( s i ) = Y 2 ( s i − ) with p r obabilit y 1 − α i , and we h av e (1 − α i ) Y 2 ( s i − ) + α i ( Y 2 ( s i − ) + 1) = Y 2 ( s i − ) + α i W AITIN G TIMES F OR MUT A TIONS 27 = Y 2 ( s i − ) 1 + 1 N − Y 1 ( s i − ) = Y 2 ( s i − ) N − Y 1 ( s i ) N − Y 1 ( s i − ) . The m artingale pr op erty follo w s b ecause A ( s i ) = A ( s i − )( N − Y 1 ( s i − )) / ( N − Y 1 ( s i )), comp ensating for the exp ected c hange in the Y 2 pro cess. Since ( Z ( t ) , t ≥ t 2 ) is a martingale conditional on ( Y 1 ( t ) , t ≥ t 1 ) and Z ( t 2 ) = 1, w e ha v e P ( Z ( t ) ≥ L/ 2 for some t | B 1 ) ≤ 2 /L . On the ev en t B 1 , we ha ve A ( t ) ≤ 2 for all t ≥ t 2 , so P ( B 2 | B 1 ) ≤ P ( Y 2 ( t ) ≥ L for some t | B 1 ) ≤ P ( Z ( t 2 ) ≥ L / 2 for some t | B 1 ) ≤ 2 /L. Since P ( B 1 ) ≤ 1 /L , the result f ollo ws. W e now int ro duce a set-up that will all o w us to apply Lemma 5.1 . Let ε > 0, and let K b e a large p ositiv e n umb er that will b e c hosen late r. Let ¯ q m b e the p robabilit y that in mo del M 1 : • there is ev en tually a typ e m individual in the p opu lation, • the maximum num b er of individuals of n on zero t yp e o v er all times is b et w een ε/r 1 ,m and N/ 2, and • the family liv es for time ≤ K/r 1 ,m ; th at is, there are no individuals of nonzero t yp e remainin g at time K/r 1 ,m . W e will call the second and third p oin ts the side c onditions . Divide the in terv al [0 , t/ ( N r 0 ,m )] in to M su bint erv als of equ al length, where M r 1 ,m → ∞ as N → ∞ . Lab el the in terv als I 1 , . . . , I M , and let D i b e the ev ent that there is a type 1 m utation in the in terv al I i . F or b o okk eeping pur p oses, we will also in tr o duce t yp e 1b mutati ons, whic h individuals of t yp e greate r than zero exp er ience at rate u 1 but which do not affect the typ e of the individual. Mutations to t yp e zero individ uals will b e called typ e 1a m u tations, and th e p hrase “t yp e 1 m utation” will refer b oth to type 1a and t yp e 1b m u tations for the rest of this section. This will ensu re that t yp e 1 mutatio ns are alw a ys o ccurring at rate exactly N u 1 . T o determine wh ether or not the first t yp e 1b mutati on in in terv al i is “successful,” we let ξ 1 , . . . , ξ M b e i.i.d. rand om v ariables, indep enden t of our pro cess, that equal 1 with p robabilit y ¯ q m . Let A i b e the eve n t that th ere is a typ e 1 mutatio n in the int erv al I i and one of the follo win g o ccurs: • The fi rst typ e 1 m utation in I i has t yp e 1a. Th e individual that gets this m u tation has a t yp e m descendant and the sid e cond itions hold. That is, the maxim um num b er of descendants of the mutatio n o ver all times 28 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG is b et w een ε/r 1 ,m and N / 2, and there are no descendants of the muta- tion remaining in the p opulation at the time K /r 1 ,m after the m u tation o ccurred. • The first t yp e 1 m u tation in I i has t yp e 1b, and ξ i = 1. As in Lemma 5.1 , let W = P M i =1 1 A i b e the n u mb er of ev ents that occur, and let λ = E [ W ]. Lemma 5.3. lim sup N →∞ | P ( W = 0) − e − λ | = 0 . Pr oof. Let β i consist of all subinte rv als whose d istance to I i is at most K/r 1 ,m . Define b 1 , b 2 and b 3 as in Lemma 5.1 . W e fi rst claim that b 3 = 0. Supp ose I i = [ a, b ]. Note that the eve n t A i do es n ot d ep end on th e state of the p opulation at time a . Also, b ecause of th e side condition that a m utation is not considered su ccessful if it has d escendan ts surviving for a time longer than K/r 1 ,m , the ev ent A i is determined b y time b + K /r 1 ,m and is therefore indep en d en t of the even ts A j for j > i and j / ∈ β i . Lik ewise, all of the ev en ts A j for j < i and j / ∈ β i are determined by the b eha vior of th e pro cess b efore time a , so these even ts are also indep enden t of A i . It follo ws that A i is indep en d en t of ( A j ) j / ∈ β i , and th us that b 3 = 0. The length | I i | of the in terv al I i is t/ ( M N r 0 ,m ), so since t yp e 1 m utations o ccur at rate N u 1 , w e ha ve P ( D i ) ≤ N u 1 | I i | = t / ( M r 1 ,m ). Since P ( A i | D i ) = ¯ q m , Prop osition 4.1 giv es P ( A i ) = ¯ q m P ( D i ) ≤ t q m / ( M r 1 ,m ) ∼ t / M . There are at most 2( K/ ( r 1 ,m | I i | ) + 1) interv als in β i , so for large M b 1 ≤ M · 2 K r 1 ,m | I i | + 1 · t M 2 = 2 M · K M N r 0 ,m r 1 ,m t t M 2 + 2 t 2 M = 2 K N u 1 t + 2 t 2 M . Since N u 1 → 0 b y (i) and M → ∞ , b 1 → 0. T o b ound b 2 , note that P ( D i ∩ D j ) ≤ [ t/ ( M r 1 ,m )] 2 b ecause mutati ons in disjoint inte r v als o ccur in dep end en tly . W e no w ap p ly Lemma 5.2 w ith L = ε/r 1 ,m , t 1 b eing the time of the first mutati on in I i , and t 2 b eing the time of the first m utation in I j . F or the ev ent A i to o ccur, it is necessary for the ev en t B i considered in Lemma 5.2 to occur. Note that mutati ons do not effect the result of Lemma 5.2 b ecause the sid e conditions in volv e all descendan ts of the original m u tation, regardless of type. P ( A i ∩ A j | D i ∩ D j ) ≤ 2 r 2 1 ,m /ε 2 W AITIN G TIMES F OR MUT A TIONS 29 and th us P ( A i ∩ A j ) ≤ 2 t 2 / ( M ε ) 2 . Since there are at most 2( K/ ( r 1 ,m | I i | ) + 1) in terv als in β i , w e ha ve b 2 ≤ M · 2 K r 1 ,m | I i | + 1 2 t 2 ( M ε ) 2 = 4 M · K M N r 0 ,m r 1 ,m t t M ε 2 + 4 t 2 M ε 2 = 4 ε − 2 K N u 1 t + 4 t 2 M ε 2 . This sho ws b 2 → 0, and completes the pr o of. Lemma 5.4. L et σ m b e the first time at which ther e is a typ e 1 individual in the p opula tion that wil l have a typ e m desc endant. Then lim N →∞ P ( σ m > t/ ( N r 0 ,m )) = exp( − t ) . (5.1) Pr oof. T o obtain ( 5.1 ) from Lemma 5.3 , it suffices to sho w that there is a constan t C such that for su fficien tly large N , we h a ve | t − λ | ≤ C ε and | P ( W = 0) − P ( σ m > t/ ( N r 0 ,m )) | ≤ C ε . The result will then follo w by letting ε → 0. Clearly , ¯ q m ≤ q m , and q m − ¯ q m is at most the p r obabilit y that in mo del M 1 , a t yp e m ind ividual app ears ev en though either (a) the total n u m b er of individu als of nonzero typ e nev er exceeds εr 1 ,m , (b) the total n u m b er of individu als of nonzero t yp e exceeds N / 2, or (c) the family do es not die out b efore K/r 1 ,m . Because N r 1 ,m → ∞ , w e ha v e K/r 1 ,m < N for sufficien tly large N , so w e can apply Lemma 3.1 to sh o w that the probabilit y that a giv en mutatio n survive s f or as long as K/r 1 ,m is at most C r 1 ,m /K . Using Lemma 4.4 , w e get q m − ¯ q m ≤ C εr 1 ,m + 2 / N + C r 1 ,m /K. Since N r 1 ,m → ∞ b y (iv), w e h a ve 2 / N ≪ r 1 ,m , s o if K is large, w e get q m − C εr 1 ,m ≤ ¯ q m ≤ q m . (5.2) Note that λ = X i ∈I P ( A i ) = X i ∈I P ( D i ) ¯ q m = M P ( D 1 ) ¯ q m = M ¯ q m (1 − e − N u 1 | I 1 | ) ∼ M ¯ q m N u 1 | I 1 | = t ¯ q m /r 1 ,m . Because q m ∼ r 1 ,m b y Prop osition 4.1 , this result com bined with ( 5.2 ) implies | t − λ | ≤ C ε for sufficien tly large N . It remains to b oun d | P ( W = 0) − P ( σ m > t/ ( N r 0 ,m )) | . W e can ha ve W > 0 with σ m > t/ ( N r 0 ,m ) only if for some i , there is a typ e 1b muta tion in I i and ξ i = 1. Let X ( t ) b e the num b er of individu als of nonzero t yp e. As long 30 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG as X ( t ) sta ys b elo w εN , t yp e 1b m u tations occur at rate at most N εu 1 , so the probabilit y that this o ccurs is at most ( εN u 1 )( t/ N r 0 ,m ) ¯ q m ≤ C ε, using Prop osition 4.1 . Since individu als give birth and die at the same rate, ( X ( t ) , t ≥ 0) is a su bmartingale. Also, E [ X ( t/ ( N r 0 ,m ))] is the exp ected n um- b er of t yp e 1a m utations b efore time t/ ( N r 0 ,m ), which is at most t/r 1 ,m . Therefore, b y Do ob ’s m aximal inequalit y , P ( X ( s ) ≥ εN for some s ≤ t/ ( N r 0 ,m )) ≤ t/ ( εN r 1 ,m ) , whic h go es to zero as N → ∞ b y cond ition (iv). W e can ha ve W = 0 with σ m ≤ t/ ( N r 0 ,m ) in one of t w o w ays. One p os- sibilit y is that there could b e a successful t yp e 1 m utation in one of the M subinterv als that is not the first type 1 m u tation in that interv al. The ex- p ected n umb er of t yp e 1 m utations in the i th in terv al that are not the fi rst in their in terv al is at most ( t/ M r 1 ,m ) 2 . Therefore, the pr obabilit y that some successful t y p e 1 m u tation is not the fi rst t yp e 1 mutatio n in its in terv al is at most M ( t/ M r 1 ,m ) 2 q m ≤ C / ( M r 1 ,m ). Since M r 1 ,m → ∞ , this probabil- it y tends to zero as N → ∞ . Th e other p ossibilit y is that there could b e a successful type 1 mutatio n that d o es not satisfy the extra conditions w e imp osed. The probabilit y that this occur s is at most ( N u 1 )( t/ N r 0 ,m )( q m − ¯ q m ) ≤ C tε b y ( 5.2 ). This observ ation completes the pro of of the lemma. The follo wing result in com bination with Lemma 5.4 implies T heorem 2 . Lemma 5.5. We have N r 0 ,m ( τ m − σ m ) → 0 in pr ob ability . (5.3) Pr oof. Let ε > 0 and δ > 0. By Lemma 5.4 , we can c h o ose s large enough that f or su fficien tly large N , P ( σ m > s / ( N r 0 ,m )) < δ / 3 . By Lemma 3.1 , the probabilit y that a typ e 1a m u tation tak es longer than time ε/ ( N r 0 ,m ) to d ie out or fixate is at most C max { 1 / N , N r 0 ,m /ε } . Be- cause the exp ected num b er of t yp e 1a m utations b efore time s/ N r 0 ,m is at most ( N u 1 )( s/ N r 0 ,m ) = u 1 s/r 0 ,m , it follo ws from Mark ov’s inequalit y that the probab ility th at some t yp e 1a muta tion that app ears b efore time s/ ( N r 0 ,m ) tak es longer th an time ε/ ( N r 0 ,m ) to d ie out or fixate is at m ost C s max { u 1 / ( N r 0 ,m ) , N u 1 /ε } . As N → ∞ , the fir st of these term s go es to zero by (iv) wh ile the second goes to zero by (i), so this probabilit y is less W AITIN G TIMES F OR MUT A TIONS 31 than δ / 3 for su fficien tly large N . Finally , the probabilit y th at one of the t yp e 1a mutatio ns b efore time s/ ( N r 0 ,m ) fixates is at most s N r 0 ,m · N u 1 · 1 N , since muta tions o ccur at rate N u 1 and fi x with p robabilit y 1 / N . Th is is less th an δ / 3 for large N b y (iv). Hence, P ( N r 0 ,m ( τ m − σ m ) > ε ) < δ for sufficien tly large N . 6. The k ey to the p ro of of Theorem 3 . Throughout this s ection and th e next, we assume all of the hyp otheses of Theorem 3 are satisfied. Th e main difficult y in proving Th eorem 3 is to prov e the follo wing result. Pr oposition 6.1. L et ε > 0 . Consider a pr o c ess which evolves ac c or ding to the rules of mo del M 1 but starting with [ εN ] typ e 1 individuals and al l other individuals having typ e 0. L et g N ,m ( ε ) b e the pr ob ability tha t either a typ e m individual is b orn at some time or eventual ly al l N individuals have typ e gr e ater than zer o. Then lim ε → 0 lim inf N →∞ ε − 1 g N ,m ( ε ) = lim ε → 0 lim sup N →∞ ε − 1 g N ,m ( ε ) = α, wher e α is as define d in ( 1.4 ). The first lemma will allo w u s to ignore o verlap b et ween t yp e 2 families. Lemma 6.1. With pr ob ability tending to one as N → ∞ , no typ e 2 in- dividual that is b orn while ther e is an individual of typ e 0 in the p opulation and another individual in the p opulation of typ e 2 or higher wil l have a typ e m desc endant. Pr oof. The argumen t is similar to the pro of of Lemma 4.3 . By ( 3.5 ), when w e start with [ N ε ] t yp e 1 ind ividuals, the total num b er of bir ths and deaths of individu als of nonzero t yp e, b efore the n u m b er of individuals of nonzero type reac hes 0 or N , is at most 2 εN 2 . S in ce individuals giv e birth and d ie at rate 1 and mutate at rate u 2 , the exp ected num b er of type 2 m u tations is at most εN 2 u 2 . By ( 3.8 ), the exp ected amoun t of time during whic h there is an individual of typ e 2 or h igher presen t in the p opulation is at m ost C ε ( N 2 log N ) u 2 . Type 2 m utations hap p en at rate at m ost N u 2 and pro duce a t yp e m d escendan t with pr ob ab ility q 2 ,m , so the probab il- it y that a type 2 individ u al b orn while there is another individu al in the p opulation of t yp e 2 or higher pro duces a t yp e m descendant is at most C εN 3 (log N ) u 2 2 q 2 ,m , w hic h is at most C ( N r 1 ,m ) 2 ( N log N ) u 2 , (6.1) 32 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG b ecause u 2 r 2 ,m = r 2 1 ,m and q 2 ,m ∼ r 2 ,m b y C orollary 4.1 . Also, we are as- suming N r 1 ,m → γ 1 / 2 , and (ii) giv es r 1 ,m ≥ C u 1 − 1 / 2 m − 1 2 for some constant C . Therefore, lim sup N →∞ N u 1 − 1 / 2 m − 1 2 < ∞ , whic h in com bination with (iii) implies that ( N log N ) u 2 → 0 . (6.2) It follo ws that the expression in ( 6.1 ) tends to zero as N → ∞ . In view of Lemma 6.1 , it suffices to pro ve Prop osition 6.1 for mo del M 2 , in which no t yp e 2 m utation can o ccur while there is another individual of t yp e 2 or higher in the p opulation. W e will work with mo del M 2 for the rest of this section. As in the pro of of Th eorem 1 , w e need to deal with the correlations b et w een ind ividuals of t yp e 1 and of t yp es j ≥ 2 caused by the fact that individuals of one p ositiv e type may r eplace another. T o do this, w e cut out the time int erv als in whic h an in dividual of t yp e 2 or higher is present in the p opulation. Let X i ( t ) b e the n umb er of t yp e i ind ividuals at time t . Let f ( t ) = sup s : Z s 0 1 { X 0 ( t )+ X 1 ( t )= N } du = t and let Y ( t ) = X 1 ( f ( t )), so the p ro cess ( Y ( t ) , t ≥ 0) trac ks the ev olution of the n u m b er of typ e 1 individuals after one cuts out the times at whic h individuals of t yp e j ≥ 2 are present. Let β 0 = 0. F or i ≥ 1, let β i b e the first time t after β i − 1 suc h th at Y ( t ) 6 = Y ( t − ) an d there is no t yp e t w o individual aliv e at time f ( t ) − , assum ing such a time exists wh ic h it will a.s. as long as Y ( β i − 1 ) / ∈ { 0 , N } . That is, the times β i are the times of Y p ro cess j umps that h app en b ecause of a birth or d eath of a t yp e one individual and do not in volv e the bir th of a t yp e tw o in dividual. Let g ( t ) = max { i : β i ≤ t } , so g ( t ) is the num b er of these jump s that ha ve happ ened b y time t . W e now d efine a discrete-time pro cess ( Z i ) ∞ i =0 , w hic h omits the jumps in Y due to time inte rv als b eing remov ed, bu t retains all of the other j umps of size 1. Let Z 0 = [ N ε ]. If i ≥ 1, Y ( β i − 1 ) / ∈ { 0 , N } , and ε 3 N < Z i − 1 < (1 − ε 2 ) N , then let Z i = Z i − 1 + 1 if Y ( β i ) = Y ( β i − ) + 1, and let Z i = Z i − 1 − 1 if Y ( β i ) = Y ( β i − ) − 1. Using this induction, we can define the p ro cess ( Z i ) T i =0 , where T = inf { i : Y ( β i ) ∈ { 0 , N } , Z i ≤ ε 3 N , or Z i ≥ (1 − ε 2 ) N } . On the ev ent that ε 3 N < Z i − 1 < (1 − ε 2 ) N and 0 < Y ( β i ) < N , w e hav e P ( Z i = Z i − 1 + 1 | Z 0 , . . . , Z i − 1 ) = P ( Z i = Z i − 1 − 1 | Z 0 , . . . , Z i − 1 ) = 1 / 2. W e then con tinue the pro cess f or i > T by setting Z i to b e Z i − 1 + 1 or Z i − 1 − 1 with probabilit y 1 / 2 eac h, ind ep end ently of the p opulation pro cess. The p ro cess ( Z i ) ∞ i =0 is therefore a simple rand om w alk. Note that T is smaller than the absorption time of the pro cess ( Z i ) ∞ i =0 in { 0 , N } , whic h can b e compared to the absorption time of mo del M 0 started W AITIN G TIMES F OR MUT A TIONS 33 with [ N ε ] t y p e 1 individ u als. It therefore follo ws from ( 3.9 ) that E [ β T ] ≤ N . Th us, if θ > 0, then by Mark ov’s inequalit y , P β T > N θ ≤ θ . (6.3) Lik ewise, since T is at most th e num b er of bir ths and deaths of individuals of nonzero t yp e started from [ N ε ] suc h individuals, ( 3.5 ) giv es E [ T ] ≤ 2 N 2 ε ≤ 2 N 2 . Therefore, for θ > 0, P T > 2 N 2 θ ≤ θ . (6.4) Lemma 6.2. F or al l δ > 0 , we have lim N →∞ P max 0 ≤ t ≤ β T | Y ( t ) − Z g ( t ) | > δ N = 0 . Pr oof. Let ζ 0 = 0 and for i ≥ 1, let ζ i b e the fi rst time t after ζ i − 1 suc h that there is a t yp e 2 individu al aliv e at time f ( t ) − , pro vid ed such a time exists. Th u s, the times ζ i for i ≥ 1 are the times at whic h the p ro cess ( Y ( t ) , t ≥ 0) p ossibly jumps b ecause w e hav e cut out the lifetime of a t yp e 2 family . E very jump time of ( Y ( t ) , t ≥ 0) is either β i or ζ i for some i . Since only the ju mps at the times β i are incorp orated into the pro cess ( Z i ) ∞ i =1 , w e ha ve Y ( t ) − Z g ( t ) = X i : ζ i ≤ t ( Y ( ζ i ) − Y ( ζ i − )) . (6.5) W e will sho w th at the righ t-hand side is small b ecause t yp e 2 ind ividuals are not aliv e in the p opulation for a long enough time f or large c h anges in the size of the t yp e 1 p opulation to hap p en du r ing this time. A type 1 individual is lost w henev er a t yp e 2 ind ividual is b orn. The other c hanges in the num b er of t yp e 1 individu als that con tribu te to th e righ t- hand side of ( 6.5 ) are b irths and deaths that o ccur while there are already t yp e 2 individu als in the p opulation. Let ξ i = 1 if the i th suc h eve n t is a birth, an d let ξ i = − 1 if the i th suc h ev ent is a death. Let J b e the n umb er of suc h ev ents b efore time f ( β T ), so if S j = ξ 1 + · · · + ξ J , th en | Y ( t ) − Z g ( t ) | ≤ |{ i : ζ i ≤ T }| + max j ≤ J | S j | (6.6) for all t ≤ β T . The fi rst term on the right-hand side of ( 6.6 ) is the num b er of t yp e 2 m u tations by time β T , so as noted ab o v e its exp ected v alue is at most εN 2 u 2 . It follo ws from Mark o v’s inequ alit y and ( 6.2 ) that P ( |{ i : ζ i ≤ T }| > δ N / 2) ≤ 4 εN 2 u 2 / ( δN ) → 0 as N → ∞ . 34 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG Since ( S j ) ∞ j =1 is a simple rand om walk, b y the monotone con vergence the- orem, the L 2 -maximal inequalit y for martin gales, and W ald’s second equa- tion, w e ha v e E max j ≤ J S 2 j = lim n →∞ E max j ≤ J ∧ n S 2 j ≤ 4 lim n →∞ E [ S 2 J ∧ n ] = 4 lim n →∞ E [ J ∧ n ] = 4 E [ J ] . W e ha ve observ ed that th e exp ected amount of time for whic h there is an individu al of t yp e 2 or greater presen t in the p opu lation is at most C ε ( N 2 log N ) u 2 . The rate at wh ich t yp e one individ u als are either b eing b orn or dying is alw a ys at most 2 N , so E [ J ] ≤ 2 C ε ( N 3 log N ) u 2 . By Ch eb y - shev’s inequalit y and ( 6.2 ), lim sup N →∞ P max j ≤ J | S j | > δ N 2 ≤ lim su p N →∞ 16 E [ J ] δ 2 N 2 ≤ lim su p N →∞ 32 C ε ( N log N ) u 2 δ 2 = 0 and the result follo ws. Lemma 6.3. F or al l δ > 0 , we have lim N →∞ P Z β T 0 ( Y ( t ) − Z g ( t ) ) dt > δ N 2 = 0 . Pr oof. Let θ > 0. By Lemma 6.2 an d ( 6.3 ), lim sup N →∞ P Z β T 0 ( Y ( t ) − Z g ( t ) ) dt > δ N 2 ≤ lim sup N →∞ P β T > N θ + P max 0 ≤ t ≤ β T | Y ( t ) − Z g ( t ) | > δ θ N ≤ θ . Letting θ → 0 giv es the result. Lemma 6.4. F or al l δ > 0 , we have lim N →∞ P Z β T 0 Z g ( t ) dt − T − 1 X i =0 N 2( N − Z i ) > δ N 2 = 0 . Pr oof. F or i ≤ T − 1 , let D i = N 2( N − Z i ) − ( β i +1 − β i ) Z i . W AITIN G TIMES F OR MUT A TIONS 35 W e n eed to sh o w that lim N →∞ P T − 1 X i =0 D i > δ N 2 ! = 0 . (6.7) A t time t , ev ents that cause th e n u m b er of t yp e 1 individuals to c hange but do not inv olv e the b irth of a t yp e 2 happ en at rate 2 Y ( t )( N − Y ( t )) / N . Therefore, if w e define ξ i = Z β i +1 β i 2 Y ( t )( N − Y ( t )) N dt, then the random v ariables ξ i are indep enden t and ha ve the exp onen tial distribution with mean one. Note th at th e pro cess Y is constan t on the in terv als ( β i , β i +1 ) except when typ e 2 m utations o ccur. F or i ≤ T − 1 , let ˜ D i = N 2( N − Z i ) (1 − ξ i ) . Let θ > 0, so P ( T > 2 N 2 /θ ) ≤ θ b y ( 6.4 ). F or 0 ≤ j ≤ [2 N 2 /θ ], let M j = P ( T − 1) ∧ j i =0 ˜ D i . Let F j b e th e σ -field generated b y ( Y ( t ) , 0 ≤ t ≤ β j ). Note that E [ ˜ D i |F i ] = 0, so the p ro cess ( M j ) [2 N 2 /θ ] j =0 is a martingale. On the ev en t that i ≤ T − 1, we ha v e Z i ≤ (1 − ε 2 ) N , and hence V ar ( ˜ D i |F i ) = N 2 4( N − Z i ) 2 ≤ 1 4 ε 4 . It follo ws from the L 2 -maximal inequalit y for martin gales, and orthogonalit y of martingale incremen ts that E max 0 ≤ j ≤ [2 N 2 /θ ] M 2 j ≤ 4 E [ M 2 [2 N 2 /θ ] ] ≤ 4 · 2 N 2 θ · 1 4 ε 4 = 2 N 2 θ ε 4 . Using Cheb yshev’s inequalit y , P T − 1 X i =0 ˜ D i > δ N 2 2 ! ≤ θ + P max 0 ≤ j ≤ [2 N 2 /θ ] | M j | > δ N 2 2 ≤ θ + 4 δ 2 N 4 2 N 2 θ ε 4 = θ + 8 θ δ 2 ε 4 N 2 . Since θ > 0 w as arbitrary , it f ollo ws that lim N →∞ P T − 1 X i =0 ˜ D i > δ N 2 2 ! = 0 . (6 .8) 36 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG T o con v ert this into a b ound on the D i , w e note that | D i − ˜ D i | = N 2( N − Z i ) Z β i +1 β i 2 Y ( t )( N − Y ( t )) N dt − ( β i +1 − β i ) Z i ≤ Z β i +1 β i Y ( t )( N − Y ( t )) N − Z i − Z i dt. On th e even t that | Y ( t ) − Z g ( t ) | ≤ γ N for all 0 ≤ t ≤ β T , th ere is a constan t C ε dep end ing on ε su c h that for all i ≤ T − 1 and t ∈ [ β i , β i +1 ], w e ha v e Y ( t )( N − Y ( t )) N − Z i − Z i ≤ ( Z i + γ N )( N − Z i + γ N ) N − Z i − Z i ≤ ( Z i + γ N ) 1 + γ ε 2 − Z i ≤ C ε γ N , where in the second in equalit y we ha v e used Z i ≤ (1 − ε 2 ) N . F or a b oun d in the other direction, w e note that Y ( t )( N − Y ( t )) N − Z i − Z i ≥ ( Z i − γ N )( N − Z i − γ N ) N − Z i − Z i ≥ ( Z i − γ N ) 1 − γ ε 2 − Z i ≥ − C ε γ N . Th us, if we let θ > 0 and γ = δ θ / 2 C ε , then for suffi cien tly large N , P T − 1 X i =0 ( D i − ˜ D i ) > δ N 2 2 ! ≤ P β T > N θ + P max 0 ≤ t ≤ β T | Y ( t ) − Z g ( t ) | > γ N . Using ( 6.3 ), Lemma 6.2 , and letting θ → 0, w e get lim N →∞ P T − 1 X i =0 ( D i − ˜ D i ) > δ N 2 2 ! = 0 . (6.9) No w ( 6.7 ) follo ws f rom ( 6.8 ) and ( 6.9 ). Let D b e the ev ent that either Z T ≥ (1 − ε 2 ) N or some t yp e 2 mutati on that occur s b efore time f ( β T ) has a t yp e m descendant. Lemma 6.5. We have lim N →∞ (1 − P ( D )) − E " exp − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) ! 1 { Z T ≤ ε 3 N } #! = 0 . W AITIN G TIMES F OR MUT A TIONS 37 Pr oof. If there is no typ e 2 individual in the p opulation at time t , then the rate at whic h a t yp e 2 individu al is b orn is u 2 X 1 ( t ). Because no t yp e 2 m u tations o ccur while th er e is another t yp e 2 individ ual in the p opu lation, eac h mutan t typ e 2 individual indep en d en tly has a t yp e m descendan t with probabilit y q 2 ,m . It follo ws that there is a mean one exp onential random v ariable ξ suc h that some original t yp e tw o individual b orn b efore time f ( β T ) has a t yp e m descendant if and only if ξ ≤ Z β T 0 Y ( t ) u 2 q 2 ,m dt. (6.10) Because c h anges in the p opulation resulting from the b irth of a t yp e 2 individual are not recorded in the pro cess ( Z i ) T − 1 i =0 , the random v ariable ξ can b e constructed to b e indep endent of the pro cess ( Z i ) T − 1 i =0 . Therefore, b y cond itioning on ( Z i ) T − 1 i =0 , we get P { Z T ≤ ε 3 N } ∩ ( ξ > r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) )! (6.11) = E " exp − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) ! 1 { Z T ≤ ε 3 N } # . The eve n t that D fails to occur is the same as the ev ent th at Z T ≤ ε 3 N and that ( 6.10 ) fails to o ccur. It follo ws that the d ifference b et wee n P ( D c ) = 1 − P ( D ) and the p r obabilit y in ( 6.11 ) is at most the p robabilit y that ξ is b et w een R β T 0 Y ( t ) u 2 q 2 ,m dt and r 2 ,m P T − 1 i =0 u 2 N/ (2( N − Z i )). T o b ound the difference b et ween these qu an tities, note that Lemmas 6.3 and 6.4 giv e lim N →∞ P Z β T 0 u 2 Y ( t ) dt − T − 1 X i =0 u 2 N 2( N − Z i ) > δ N 2 u 2 ! = 0 for all δ > 0. S ince r 2 1 ,m = u 2 r 2 ,m and ( N r 1 ,m ) 2 → γ , w e see that N 2 u 2 r 2 ,m sta ys b ounded as N → ∞ and it follo ws that lim N →∞ P Z β T 0 u 2 r 2 ,m Y ( t ) dt − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) > δ 2 ! = 0 (6.12) for all δ > 0. Also, q 2 ,m ∼ r 2 ,m b y Corollary 4.1 and P ( β T > N /θ ) ≤ θ b y ( 6.3 ). Since N 2 u 2 r 2 ,m sta ys b ounded, lim sup N →∞ P Z β T 0 u 2 q 2 ,m Y ( t ) dt − Z β T 0 u 2 r 2 ,m Y ( t ) dt > δ 2 (6.13) ≤ lim sup N →∞ P N u 2 β T | r 2 ,m − q 2 ,m | > δ 2 = 0 . 38 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG Com b ining ( 6.1 2 ) and ( 6.13 ) giv es lim N →∞ P Z β T 0 u 2 q 2 ,m Y ( t ) dt − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) > δ ! = 0 . Since P r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) − δ ≤ ξ ≤ r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) + δ ! ≤ 2 δ , it follo ws that lim sup N →∞ (1 − P ( D )) − E " exp − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) ! 1 { Z T ≤ ε 3 N } # ≤ 2 δ, and the result follo ws b y letting δ → 0. Let A b e the ev ent that either Y ( t ) = N for some t , or a t yp e m individu al is b orn at s ome time. Lemma 6.6. Ther e exists a c onstant C , not dep ending on ε or N , such that | P ( A ) − P ( D ) | ≤ C ε 2 . Pr oof. Let δ > 0 , and assume that | Y ( t ) − Z g ( t ) | ≤ δ N for 0 ≤ t ≤ β T . First, supp ose D occur s . If a t yp e 2 m u tation that o ccurs b efore time f ( β T ) has a type m descendant, then A m ust occur. If Z T ≥ (1 − ε 2 ) N , then Y ( β T ) ≥ (1 − ε 2 − δ ) N , and conditional on this eve n t the probabilit y that Y ( t ) = N for some t , in which case A o ccurs, is at least 1 − ε 2 − δ . Th er efore, using Lemma 6.2 , lim sup N →∞ P ( D ∩ A c ) ≤ ε 2 + δ. No w, supp ose D c o ccurs. Note that if δ < ε 3 and | Y ( t ) − Z g ( t ) | ≤ δ N for 0 ≤ t ≤ β T , then we cannot ha v e Y ( β T ) ∈ { 0 , N } , whic h m eans w e m ust hav e Z T ≤ ε 3 N and, therefore, Y ( β T ) ≤ ( ε 3 + δ ) N . Con d itional on this ev ent, th e probabilit y that Y ( t ) = N for some t is at most ε 3 + δ , and the probabilit y that on e of the t yp e one individuals at time f ( β T ) has a type m d escendan t is at most ( ε 3 + δ ) N q 1 ,m . F r om these b oun ds and Lemma 6.2 , it follo ws that lim sup N →∞ P ( D c ∩ A ) ≤ (1 + γ 1 / 2 )( ε 3 + δ ) . The lemma f ollo ws by letting δ → 0. No w let ( B t ) t ≥ 0 b e a Bro wnian motion with B 0 = ε . Let U = inf { t : B t = ε 3 or B t = 1 − ε 2 } . W AITIN G TIMES F OR MUT A TIONS 39 Lemma 6.7. We have lim N →∞ E " exp − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) ! 1 { Z T ≤ ε 3 N } # = E exp − γ 2 Z U 0 1 1 − B t dt 1 { B U = ε 3 } . Pr oof. Define a pr o cess ( W t ) t ≥ 0 suc h that W t = N − 1 Z [ N 2 t ] . Let R = inf { t : W t ≤ ε 3 or W t > 1 − ε 2 } . Note that R = T / N 2 and 1 { Z T ≤ ε 3 N } = 1 { W R ≤ ε 3 } on th e even t that for some δ < ε 3 , we h av e | Y ( t ) − Z g ( t ) | ≤ δ N for 0 ≤ t ≤ β T , whic h by L emm a 6.2 happ ens with pr obabilit y tending to one as N → ∞ . Let δ < ε 3 . F or ran d om v ariables X (1) N and X (2) N , write X (1) N ≈ X (2) N if for all η > 0, there is an N ( η ) su c h that if N ≥ N ( η ) then | X (1) N /X (2) N − 1 | < η on the even t that | Y ( t ) − Z g ( t ) | ≤ δ N for 0 ≤ t ≤ β T . W e hav e 1 2 Z R 0 1 1 − W t dt ≈ 1 2 Z R 0 1 1 − N − 1 Z [ N 2 t ] dt = 1 2 Z N 2 R 0 1 1 − N − 1 Z [ s ] N − 2 ds = N − 2 Z T 0 N 2( N − Z [ s ] ) ds = N − 2 T − 1 X i =0 N 2( N − Z i ) . Since u 2 r 2 ,m = r 2 1 ,m and ( N r 1 ,m ) 2 → γ , w e ha v e r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) ≈ γ N − 2 T − 1 X i =0 N 2( N − Z i ) ≈ γ 2 Z R 0 1 1 − W t dt. In view of Lemma 6.2 , it follo ws that lim N →∞ E " exp − r 2 ,m T − 1 X i =0 u 2 N 2( N − Z i ) ! 1 { Z T ≤ ε 3 N } # − E exp − γ 2 Z R 0 1 1 − W t dt 1 { W R = ε 3 } ! = 0 . Th us, it s uffices to sh o w that for all λ > 0, we hav e lim N →∞ E exp − λ Z R 0 1 1 − W t dt 1 { W R = ε 3 } (6.14) = E exp − λ Z U 0 1 1 − B t dt 1 { B U = ε 3 } . Since ( Z i ) ∞ i =0 is a simple random wa lk, ( W t ) 0 ≤ t ≤ s con verge s w eakly as N → ∞ to ( B t ) 0 ≤ t ≤ s for all s > 0. Let D [0 , s ] b e the set of real-v alued func- tions defined on [0 , s ] w h ic h are r ight conti n u ous and h a ve left limits. If 40 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG g : D [0 , s ] → R is b ound ed, and if the set of p oint s at wh ic h it is n ot con tin- uous h as Wiener measure zero, then the we ak conv ergence of ( W t ) 0 ≤ t ≤ s to ( B t ) 0 ≤ t ≤ s implies that lim N →∞ E [ g (( W t ) 0 ≤ t ≤ s )] = E [ g (( B t ) 0 ≤ t ≤ s )] . Th er e- fore, lim N →∞ E exp − λ Z R ∧ s 0 1 1 − W t dt 1 { W R ∧ s = ε 3 } (6.15) = E exp − λ Z U ∧ s 0 1 1 − B t dt 1 { B U ∧ s = ε 3 } . Note that if ω : [0 , s ] → R is con tinuous, then the function g used in ( 6.15 ) is con tinuous at ω unless either inf { t : ω ( t ) = ε 3 } < in f { t : ω ( t ) < ε 3 } or inf { t : ω ( t ) = 1 − ε 2 } < in f { t : ω ( t ) > 1 − ε 2 } , which w ould happ en if ω reac hes a lo cal min- im u m when it first hits ε 3 or a lo cal maximum wh en it first h its 1 − ε 2 . Bro wnian motio n paths almost su rely d o not hav e this prop ert y , so ( 6.15 ) is v alid. Finally , ( 6.14 ) follo ws from ( 6.15 ) b y letting s → ∞ . Let V = inf { t : B t = 0 or B t = 1 } . Lemma 6.8. L et I ( s ) = R s 0 1 1 − B t dt . If λ > 0 , ther e is a c onstant C such that | E [exp( − λI ( U )) 1 { B U = ε 3 } ] − E [ exp( − λI ( V )) 1 { B V =0 } ] | ≤ C ε 2 . (6.16) Pr oof. Define a p r o cess ( B ′ t ) t ≥ 0 b y B ′ t = B U + t . Let τ ′ a = inf { t : B ′ t = a } . Let D 1 b e the even t that B U = 1 − ε 2 and B V = 0. Let D 2 b e the even t that B U = ε 3 and τ ′ 1 / 2 < τ ′ 0 . Let D 3 b e the even t that B U = ε 3 and τ ′ 0 > ε 2 . Note that on the ev ent ( D 1 ∪ D 2 ∪ D 3 ) c , we hav e 1 { B U = ε 3 } = 1 { B V =0 } and on this ev ent w e ha ve 0 ≤ Z V 0 1 1 − B t dt − Z U 0 1 1 − B t dt ≤ 2( V − U ) ≤ 2 ε 2 . It follo w s that the left-hand side of ( 6.16 ) is at most P ( D 1 ) + P ( D 2 ) + P ( D 3 ) + 2 λε 2 . Because Bro wnian motio n is a martingale, we hav e P ( D 1 ) ≤ P ( B V = 0 | B U = 1 − ε 2 ) = ε 2 and lik ewise P ( D 2 ) ≤ 2 ε 3 . Therefore, it remains only to sho w th at P ( D 3 ) ≤ C ε 2 . By the reflection principle, 1 2 P ( τ ′ 0 ≤ ε 2 | B U = ε 3 ) = P ( B ′ ε 2 ≤ 0) . Also, P ( B ′ ε 2 > ε 3 | B U = ε 3 ) = 1 / 2. Therefore, P (0 < B ′ ε 2 < ε 3 | B U = ε 3 ) = [1 − P ( τ ′ 0 ≤ ε 2 | B U = ε 3 )] / 2. It follo ws that P ( D 3 ) ≤ P ( τ ′ 0 > ε 2 | B U = ε 3 ) = 2 P (0 < B ′ ε 2 < ε 3 | B U = ε 3 ) ≤ 2 ε 3 √ 2 π ε 2 = ε 2 r 2 π W AITIN G TIMES F OR MUT A TIONS 41 and the result follo ws. Lemma 6.9. L et E x denote e xp e ctation for the Br ownian motion ( B t ) t ≥ 0 starting fr om B 0 = x . L et u ( x ) = E x exp − γ 2 Z V 0 1 1 − B t dt 1 { B V =0 } . Then lim x → 0 x − 1 (1 − u ( x )) = α , wher e α is as define d in ( 1.4 ). Pr oof. W e c h o ose f so that f (0) = 1 and f (1) = 0. Let g ( x ) = γ / [2(1 − x )]. T hen for 0 < x < 1, w e ha ve u ( x ) = E x [ f ( B V ) exp( − R V 0 g ( B s ) ds )]. Clearly u (0) = 1 and u (1) = 0. By the F eynman–Kac form u la (see (6.3 ) on page 161 of [ 6 ]), if v : [0 , 1] → R is a b ounded con tinuous function such that v (0) = 1, v (1) = 0 , and 1 2 v ′′ ( x ) − g ( x ) v ( x ) = 0 for x ∈ (0 , 1), then u ( x ) = v ( x ) for x ∈ [0 , 1]. Note that (6.3) on page 161 of [ 6 ] requires g to b e b ounded on (0 , 1), whic h it is not in this example. Ho wev er, the result nevertheless holds b ecause g is nonn egativ e and, therefore, exp( − R t 0 g ( B s ) ds ) is alw a ys in [0 , 1]. Multiplying b y 2(1 − x ), we can write the differen tial equation ab o ve as (1 − x ) v ′′ ( x ) − γ v ( x ) = 0. Let v ( x ) = c ∞ X k =1 γ k k !( k − 1)! (1 − x ) k , (6.17) where c = 1 / P ∞ k =1 γ k /k !( k − 1)!. Note th at v (0) = 1 and v (1) = 0. The series con verge s absolutely and uniformly on all compact subsets of R and can b e differen tiated t wice term b y term, so (1 − x ) v ′′ ( x ) = c ∞ X k =2 γ k k !( k − 1)! k ( k − 1)(1 − x ) k − 1 . Therefore, (1 − x ) v ′′ ( x ) − γ v ( x ) = c ∞ X k =1 γ k +1 k !( k − 1)! (1 − x ) k − γ k +1 k !( k − 1)! (1 − x ) k = 0 . Th us, v ( x ) = u ( x ) for x ∈ [0 , 1]. F rom our form ula, it follo ws that lim x → 0 1 − u ( x ) x = − u ′ (0) = c ∞ X k =1 γ k ( k − 1)!( k − 1)! = α, as claimed. Pr oof of Proposition 6.1 . The only difference b et ween g N ,j ( ε ) and P ( A ) is that the ev ent A is defined using m o del M 2 , in whic h new t yp e t wo ind ividuals cannot b e b orn while there is an existing individual of t yp e 42 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG 2 or h igher in th e p opulation. Therefore, it follo w s from Lemma 4.3 th at | P ( A ) − g N ,j ( ε ) | ≪ [ N ε ] r 1 ,m and, therefore, lim N →∞ | P ( A ) − g N ,j ( ε ) | = 0 for all ε > 0 . By Lemm as 6.5 , 6.6 , 6.7 and 6.8 , lim sup N →∞ | P ( A ) − (1 − u ( ε )) | ≤ C ε 2 . Com b ining these r esults and multiplying b oth sides by ε − 1 giv es lim sup N →∞ | ε − 1 g N ,m ( ε ) − ε − 1 (1 − u ( ε )) | ≤ C ε. Therefore, b y L emma 6.9 , lim ε → 0 lim inf N →∞ ε − 1 g N ,m ( ε ) ≥ lim ε → 0 ( ε − 1 (1 − u ( ε )) − C ε ) = α, lim ε → 0 lim sup N →∞ ε − 1 g N ,m ( ε ) ≤ lim ε → 0 ( ε − 1 (1 − u ( ε )) + C ε ) = α and the prop osition follo ws. 7. Pro of of Theorem 3 . Wit h Prop osition 6.1 established, the rest of the pro of is routine. Lemma 7.1. Consider mo del M 1 , and let q ′ m b e the pr ob ability that either a typ e m individual is b orn at some time, or at some time al l individuals in the p opulation have typ e gr e ater than zer o. Then lim N →∞ N q ′ m = α . Pr oof. The probabilit y that the num b er of individuals of t yp e greater than zero reac h es [ εN ] is 1 / [ εN ]. If, at the time T when the num b er of individuals of nonzero t yp e reac hes [ εN ] , w e c hange th e t yp e of all these individuals to typ e 1, then the probabilit y of either getti ng a typ e m indi- vidual or ev entually ha ving all N individ u als of t yp e greater than zero is g N ,m ( ε ). Since changing the typ es in this w ay can only reduce the pr ob ab ility of in terest, we ha v e q ′ m ≥ 1 [ εN ] g N ,m ( ε ) . T o get an up p er b ound, note that the p robabilit y of either ha vin g a t yp e m ind ividual that is descend ed from a t yp e 1 ind ividual at time T or h aving all N individuals of nonzero t yp e is at most g N ,m ( ε ) / [ εN ]. The only p ossi- bilit y not acco unt ed for is that the t yp e m individual could b e descended from a t yp e 2 individual th at is b orn b efore time T . Ho we v er, by Lemma 4.4 , the pro of of which is v alid un d er our hypotheses b y Corollary 4.1 , the probabilit y that a t yp e 2 m u tation that o ccurs while there are few er than W AITIN G TIMES F OR MUT A TIONS 43 εr − 1 1 ,m individuals in the p opulation of t yp e 1 or higher has a t yp e m de- scendan t is at most C ε/ N , where we are using that r 1 ,m is O ( N ) . It follo ws that q ′ m ≤ 1 [ εN ] g N ,m ( ε ) + C ε N . The r esu lt f ollo ws from Prop osition 6.1 by first letting N → ∞ and then letting ε → 0. Pr oof of Theorem 3 . As in the p ro of of Theorem 2 , call ordinary t yp e 1 m utations t yp e 1a, and giv e eac h individual of type greater than zero a t yp e 1b m u tation at rate u 1 . Mutations of t yp e 1a and 1b will b oth b e called type 1 m utations. Let γ i b e th e time of the i th t yp e 1 m utation, so the p oints ( γ i ) ∞ i =1 form a rate N u 1 P oisson pro cess on [0 , ∞ ) . Define a sequence ( ζ i ) ∞ i =1 suc h that ζ i = 1 if the m u tation at time γ i is a t yp e 1a m u tation and has a t yp e m descendant in the p opu lation at some later time (whic h will alwa ys happ en if the m u tation fixates). Let ( ˜ ζ i ) ∞ i =1 b e a sequence of i.i.d. rand om v ariables, indep en den t of the p opulation p ro cess, suc h that P ( ˜ ζ i = 1) = q ′ m and P ( ˜ ζ i = 0) = 1 − q ′ m for all i . Let ζ ′ i = ζ i if all individuals at time γ i − ha ve type 0, and let ζ ′ i = ˜ ζ i otherwise. Let σ ′ m = inf { γ i : ζ ′ i = 1 } . It is clear from the construction that σ ′ m has th e exp onen tial distribu tion with rate N u 1 q ′ m , s o Lemma 7.1 giv es lim N →∞ P ( u 1 σ ′ m > t ) = exp ( − αt ) . (7.1) Let σ m = in f { γ i : ζ i = 1 } , which is the first time at whic h a typ e 1a m uta- tion o ccur s and the individual th at gets this mutation will ev en tu ally ha ve a t yp e m descendant. W e claim that P ( σ ′ m = σ m ) → 1 as N → ∞ . W e can only ha v e σ ′ m 6 = σ m if there is a type 1 m utation at some time γ i ≤ σ ′ m suc h that not all m u tations at time γ i − ha ve t yp e 0 and either ζ i = 1 or ˜ ζ i = 1. Note also that in this case the first suc h γ i m u st o ccur b efore any typ e 1 mutatio n fixates, so it suffices to consider the γ i that o ccur b efore an y fixation. Fix t > 0. The expected n umb er of t yp e 1 mutations b efore time u − 1 1 t is ( N u 1 )( u − 1 1 t ) = N t , so by ( 3.8 ), the exp ected amount of time b efore u − 1 1 t and b efore any typ e 1 muta tion fixates that there is an ind ividual of nonzero t yp e in the p opulation is at most C ( N log N ) t . Therefore, the ex- p ected num b er of t yp e 1 mutations that o ccur b efore this time is at most C ( N 2 log N ) u 1 t . If suc h a birth o ccurs at time γ i , the probability that either ζ i or ˜ ζ i equals one is at most 2 q ′ m , s o P ( σ m 6 = σ ′ m < u − 1 t ) ≤ C ( N 2 log N ) u 1 tq ′ m → 0 , where w e are u sing that u 1 ( N log N ) → 0 b y (ii) and ( 6.2 ) and that q ′ m is O (1 / N ) b y Lemma 7.1 . The fact that P ( σ ′ m = σ m ) → 1 as N → ∞ follo w s from this r esult and ( 7.1 ). 44 R. DURRETT, D. SCHMIDT AN D J. S CHW EINSBERG It remains only to sho w that u 1 ( τ m − σ m ) → p 0. When the t yp e 1 m u tation at time σ m do es not fi xate, τ m − σ m is at most the time that it tak es b efore all descendants of the mutatio n die out. When this m utation fi xates, then τ m − σ m includes b oth the time to fixation plus the time for one individ ual to get m − 1 additional muta tions. The probabilit y that a giv en t yp e 1 mutat ion tak es time εu − 1 1 to fixate or die out is at most C u 1 ε − 1 log N , so th e p r obabil- it y that some mutat ion that o ccurs b efore time u − 1 1 t tak es this long to fixate or die out is at most C ( N u 1 )( u − 1 1 t )( u 1 ε − 1 log N ), whic h approac h es zero as N → ∞ b ecause u 1 ( N log N ) → 0. Finally , if a t yp e 1 m u tation fixates, then the time until a t yp e m mutatio n app ears can b e calculated usin g the m − 1 case of Theorem 2 with u 2 , . . . , u m in place of u 1 , . . . , u m − 1 . T he hypotheses are satisfied b y the argu m en ts giv en in Corollary 4.1 . Th eorem 2 implies that the wa iting time is O (1 / ( N u 2 r 2 ,m )). Ho wev er, 1 / ( N u 2 r 2 ,m ) ≪ u − 1 1 b ecause u 1 /u 2 < b − 1 1 b y (ii) and N r 2 ,m → ∞ as sho wn in the p ro of of Corollary 4.1 . These observ ations imply u 1 ( τ m − σ m ) → p 0, as in the pro of of Theorem 2 . Ac kn o wledgment s. Th e authors w ould lik e to thank a r eferee whose care- ful r eading of th e pap er and m an y detailed comment s impro ve d the presen- tation of the pap er. REFERENCES [1] Armit age, P. and Doll, R. (1954). The age d istribution of cancer and a multi-stage theory of carcinogenesis. Brit. J. Canc er 8 1–12. [2] Arra tia, R . , Goldstein, L. and Gordon, L. (1989). Two moments suffice for Pois- son appro ximations: The Chen–Stein meth od . A nn. Pr ob ab. 17 9–25. MR972770 [3] A threy a, K. B. and Ne y, P. E. (1972). Br anching Pr o c esses . Springer, New Y ork. MR0373040 [4] Bor odin, A. N. and Salminen , P. (2002). Handb o ok of Br ownian Motion: F acts and F ormulae , 2nd ed. Birkh¨ auser, Boston. MR1912205 [5] Calabrese, P. , Mecklin, J. P. , J ¨ ar vinen, H. J. , Aal tonen, L. A. , T a v ar ´ e, S. and S hiba t a, D. (2005). Numbers of m utations to different typ es of colorectal cancer. BMC Canc er 5 126. [6] Durrett, R . (1996). Sto chastic Calculus: A Pr actic al Intr o duction . CRC Press, Boca Raton, FL. MR1398879 [7] Durrett, R . (2005). Pr ob ability: The ory and Examples , 3rd ed. Duxbury , Belmont, CA. MR1609153 [8] Durrett, R . and Schmidt, D. (2007). W aiting for regulatory sequences to app ear. Ann . Appl. Pr ob ab. 17 1–32. MR2292578 [9] Durrett, R . and Schmidt, D. (2008). W aiting for tw o mutations: With ap p lications to regulatory sequen ce evolution and the limits of Darwinian evol ution. Genetics 180 1501–150 9. [10] Ethier, S. N. and Kur tz, T. G . (1986). Markov Pr o c esses: Char acter ization and Conver genc e . W iley , New Y ork. MR838085 [11] Ewens, W. J. ( 2004). Mathematic al Population Genetics , 2nd ed. Springer, Berlin. MR554616 W AITIN G TIMES F OR MUT A TIONS 45 [12] Iw asa, Y. , Michor, F. , Komar ov a, N. L. and Now ak, M. A. (2005). Population genetics of tumor supp ressor genes. J. The or et. Biol. 233 15–23. MR2122451 [13] Iw asa, Y. , Michor, F. and No w ak, M. A. (2004). Stochastic tunnels in ev olutionary dynamics. Genetics 166 1571–1579. [14] Jones, S . (2008). Comparative lesion sequencing p ro vides insights into tu mor evolu- tion. Pr o c. Natl. A c ad. Sci. USA 105 4283–428 8. [15] Knudson, A. G. (1971). Mutation and cancer: Statistical study of retinoblastoma. Pr o c. Natl. A c ad. Sci. USA 68 820–823. [16] Kol mor ogo v, A. N. (1938). Zur L¨ osung einer biologisc hen Aufgabe. Izv. NI I Mat. Mekh. T omsk. Univ. 2 1–6. [17] Komar ov a, N. L. , Sengupt a, A. and No w a k, M. A. (2003). Mutation-selection netw orks of cancer initiation: T umor suppressor genes and chromosomal insta- bilit y . J. The or et. Biol. 223 433–45 0. MR2067856 [18] Luebeck, E. G . and Moolga vkar, S. H . (2002). Multistage carcinogenesis and the incidence of colorectal cancer. Pr o c. Natl. A c ad. Sci. 99 15095–15100. [19] Moran, P. A . P. (1958). Random pro cesses in genetics. Pr o c. Cambridge Philos. So c. 54 60–71. MR0127989 [20] Now ak, M. A . (2006). Evolutionary Dynamics: Exploring the Equations of Life . Belknap Press, Cambridge, MA. MR2252879 [21] Schinaz i, R. B . (2006). The probabilit y of treatment indu ced drug resistance. A cta Biothe or etic a 54 13–19. [22] Schinaz i, R. B . (2006). A sto chastic model for cancer risk. Genetics 174 545–547. [23] Schweinsberg, J. (2008). The waiti ng time for m mutations. Ele ctr on. J. Pr ob ab. 13 1442–1478 . MR2438813 [24] Wod arz, D. and Komar ov a, N. L. (2005 ). Computational Biolo gy Of Canc er: L e c- tur e Notes And Mathematic al Mo deling . W orld Scientific, S ingapore. R. Durrett Dep ar tment of Ma thema tics 310 Malott Hall Cornell University Ithaca, New York 14 8 53-4201 USA E-mail: rtd1@cornell.edu D. Schmidt Institute for Ma thema tics and its Applica tions 114 Lind Hall, 207 Chu rch St SE University of Minnesot a Minneapolis, Minnesot a 55455 USA E-mail: dsc hmidt@ima.umn.edu J. Schweinsberg Dep ar tment of Ma thema tics, 0112 University of California, San Diego 9500 Gilman Drive La Jolla, California 9209 3-0112 USA E-mail: jsch wein@math.uc sd.edu
Original Paper
Loading high-quality paper...
Comments & Academic Discussion
Loading comments...
Leave a Comment