Soliton-like base pair opening in a helicoidal DNA: An analogy with helimagnet and cholesterics
We propose a model for DNA dynamics by introducing the helical structure through twist deformation in analogy with the structure of helimagnet and cholesteric liquid crystal system. The dynamics in this case is found to be governed by the completely …
Authors: ** M. D., V. V. **
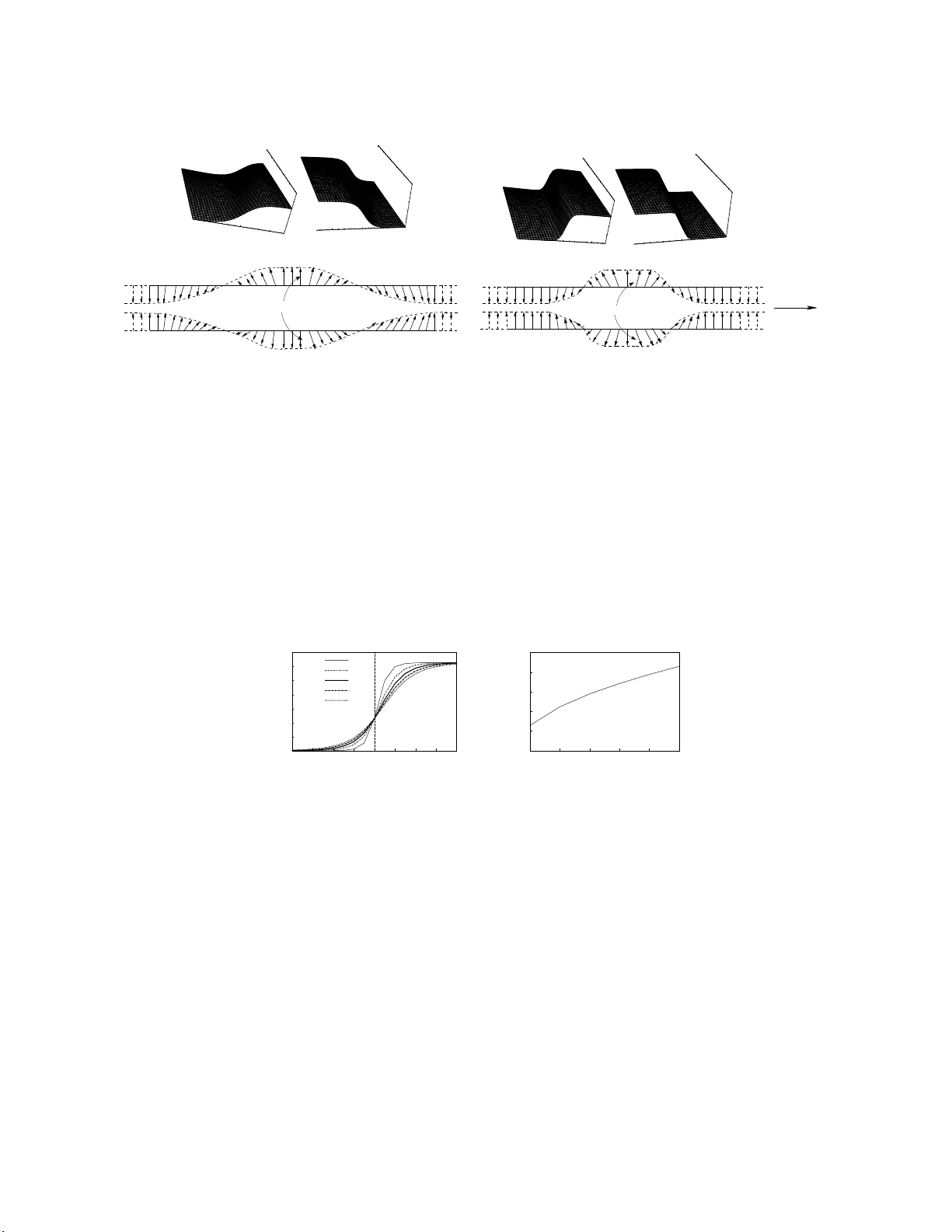
Soliton-lik e base pair op ening in a helicoidal DNA : An analogy with helimagnet and c holesterics M. Daniel ∗ and V. V asumathi † Centr e for Nonline ar D ynamics, Scho ol of Physics, Bhar athidasan University, Tiruchir app al li - 620 024, India. (Dated: Marc h 11, 2022) Abstract W e prop ose a mo del for DNA dynamics b y introd ucing the helical structur e th rough twist de- formation in a nalogy with the structure o f helimagnet and c holesteric liquid crystal system. The dynamics in this case is foun d to b e go ve rned by the complete ly integ rable sine-Gordon equation whic h admits kink-an tikink solitons with increased width represent ing a wide b a se pair op ening configuration in DNA. Th e r esu lt s sho w that the helicit y introd uces a length scale v ariation and th us provides a b ett er r ep resen tation of the base pair open in g in DNA. P ACS nu mbers: 87.1 4 .Gg, 05.45.Yv, 02.30 .Jr ∗ Electronic addres s: daniel@cnld.b du.ac.in † Electronic a ddress: v asu@cnld.b du.ac.in 1 The B-fo rm D NA double helix mo lecule is usually mo deled by t w o parallel c hains of n ucleotides kno wn a s strands with link age in terms of dip ole-dip ole inte raction along the strands a nd the tw o strands a r e coupled to eac h other t hr o ugh h ydrogen b onds b et w een the complemen tary bases [1]. Molecular excitations in DNA based on t he ab o v e mo del is generally go v erned by nonlinear ev olution equations [2, 3, 4] and in particular by the completely in t egr a ble sine-Gordon-type equations [5, 6]. In t he ab o v e studies, D NA is treated a s tw o coupled linear c hains without in v olving the helical c haracter of its structure. Ho w ev er, in nat ure DNA exists in a double helix for m and recen tly there w ere attempts b y few a ut ho rs to study the dynamics b y taking in to account the helical c haracter o f the double helix through different forms of coupling. F or instance, Gaeta [7, 8, 9], Dauxios [10] and Cadoni et al [11 ] assumed that the torsional coupling b et w een the n th base on one strand and the ( n + 4) th base on the complemen tary strand is the resp onsible force for the helical nature in DNA and f o und that the lo calized excitations ar e go v erned by solitons and breathers. Barbi et al [12, 13 ] and Campa [14] how ev er in tro duced the helicit y through a prop e r c hoice of the coupling b e t w een the radial and the angular v ariables of the helix and obta ine d breathers and kinks . On the other hand, v ery recen tly , T ak eno [15] in tr o duced helicity in DNA thro ug h a helical transformation and obtained non-breathing compacton-lik e mo des to re presen t base pair op ening through n umerical calculations. In this, paper, w e prop ose a mo del b y in tro ducing the helical character in eac h strand of the DNA molecule through a twis t deformation of the c hain in analogy with the tw ist in c holesteric liquid crystal [16] or orientation o f spins in a helimagnet [17]. As an illus tration in Fig. 1(a-c) we hav e presen ted a sc hematic represen tation of the a r rangeme n t of bases, spins and molecules resp ectiv ely in a DNA double helical chain, in a helimagnet and in a c holesteric liquid crystal leading to the formation of helical structure. In Fig. 1(a) R and R ′ represen t the t w o c omplemen tary strands of the D NA double helix and the dots b et w een the arrow s represen t the hydrogen b onds b et w een the complemen tary bases. The arrows and short lines in Figs. 1(b) and 1(c) resp ective ly represen t the spins and molecules at differen t sites and planes in a helimagnet and in a c holesteric liquid crystal. When w e go along the z-direction, the or ie n tation of spins and molecules are tilted f rom one plane to the next through certain tilt angle. If w e join the tips of the arrow s represen ting the spin v ectors and also, the tips of the molecules they form a helix as show n in Figs. 1(b) and 1(c) resp e ctiv ely . 2 z x y k ^ (a) (b) (c) z ^ x y k | R R z k ^ FIG. 1: A s c hematic r epresen tation of (a) a DNA double helical c hain (b) a helimagnet a nd (c ) a c h olesteric liquid crystal system In a recen t pap er, one of the presen t authors studied the nonlinear spin dynamics of a helimagnet by incorp orating the helicity in t erms of F rank free energy corresponding to the t wist deformation whic h is res p onsible for helicit y in a c holesteric liquid crystal system [17, 1 8]. The F ra nk free energy densit y asso ciated with the twist deformation in a c ho lesteric liquid crystal is giv en b y [ p · ( ∇ × p ) − q 0 ] 2 where t he unit v ector p repres en ts the director axis whic h corr esp onds t o the a v era g e direction of orientation of the liquid crystal molecules, q 0 = 2 π q is the pitc h w a v e v ector and q is the pitc h of the helix . The discre tised form of the ab o ve t wist free energy is written as { [ ˆ k · ( p n × p n + 1 )] − q 0 } 2 where ˆ k is the unit v ector along z-direction. In analogy with the ab o ve, we write do wn the free energy asso ciated with the t wist deformatio n in terms of spin vec tor as { [ ˆ k · ( S n × S n + 1 )] − q 0 } 2 . By taking into account the f orm of free energy the Heisen b erg mo del of Hamiltonian for an anisotro pic helimagnetic system is written as [17] H 1 = X n − J ( S n · S n + 1 ) + A ( S n z ) 2 + h { [ ˆ k · ( S n × S n + 1 )] − q 0 } 2 i . (1) In Eq.(1), S n = ( S x n , S y n , S z n ) represen ts the spin vec tor at the n th site and the terms prop or- tional t o J and A resp ectiv ely represen t the ferromagnetic spin-spin exc hange in teraction and uniaxial magneto-crystalline anisotropy with t he easy axis along z-direction. h de- 3 notes the elastic constan t asso ciated with the t wist deformatio n. W e iden tify the ab o v e helical spin c hain with one of the strands of the DNA double helical ch ain. Therefore, in a similar fashion w e can write dow n the spin Hamiltonian H 2 for another helimagnetic system corresp onding to the complemen tary strand with the spin v ector S n replaced b y S ′ n . W e assume that in the Hamiltonian the exc ha nge, anisotropic and twist co efficien ts as w ell as the pitc h in b oth the helimagnetic systems ar e equal. Now, f or mapping the helimagnetic spin system with the DNA double helical c hain we rewrite the Hamilto nian b y writing the spin v ectors as S n ≡ ( S x n , S y n , S z n ) = (sin θ n cos φ n , sin θ n sin φ n , cos θ n ) and S ′ n ≡ ( S ′ x n , S ′ y n , S ′ z n ) = (sin θ ′ n cos φ ′ n , sin θ ′ n sin φ ′ n , cos θ ′ n ), where θ n ( θ ′ n ) and φ n ( φ ′ n ) ar e the angles of rotatio n of spin s in the xy and xz-planes r e sp ectiv ely . The new Ha milto nian cor- resp onding to H 1 is written as H 1 = X n [ − J { sin θ n sin θ n +1 cos( φ n +1 − φ n ) + cos θ n cos θ n +1 } + A cos θ 2 n + h { sin θ n × sin θ n +1 sin( φ n +1 − φ n ) − q 0 } 2 , (2) W e n o w m ap the t w o helical spin systems with the tw o s trands of the DNA double helix with the t w o angles θ n ( θ ′ n ) and φ n ( φ ′ n ) represen ting the angles of rotatio n of ba ses in the xz and xy-planes of the t w o strands resp ec tiv ely . A horizontal pro jection of the n th base of DNA in the xy and xz-planes is shown in Figs. 2(a) and 2(b). Here Q n and Q ′ n denote the tips of the n th bases attac hed to the strands R and R ′ at P n and P ′ n resp e ctiv ely . The DNA double helix chain is stabilised by stac king of ba ses through in trastrand dip ole-dip ole in teraction and through h ydrogen b onds (in terstrand interaction) betw een complemen tary bases. The in terstrand base-base interaction or h ydrogen b o ndin g energy betw een the compleme n tary bases dep end s on the distance b et w een them and using the simple geometry in F ig s . 2(a,b), w e can write the distance b et w een the t ips of bases a s [6], ( Q n Q ′ n ) 2 ≈ 2 [sin θ n sin θ ′ n (cos φ n cos φ ′ n + sin φ n sin φ ′ n ) − cos θ n cos θ ′ n ] . (3) No w, the ab o v e equation represe n ts the h ydrogen bonding energy betw een complemen tary bases and w e can write do wn the Ha milto nian for the inters trand in teraction o r hydrogen 4 | | | y x O n n P φ n φ n n n P Q Q | | | | | O P n θ n n Q n Z n Z n Q θ n n P X Z (a) (b) FIG. 2: A horizon tal pro jection of the n th base pair in a DNA double helix (a) in the x y -p la ne and (b) in the xz -plane. b onds is written as H 12 = η [sin θ n sin θ ′ n (cos φ n cos φ ′ n + sin φ n sin φ ′ n ) − cos θ n cos θ ′ n ] , (4) where η is a constant. The total Hamiltonian for our helicoidal mo del of DNA in terms of the angles of rotation of bases using the ab ov e Hamiltonians is written as H = H 1 + H 2 + H 12 = X n [ − J { sin θ n sin θ n +1 cos( φ n +1 − φ n ) + cos θ n × cos θ n +1 + sin θ ′ n sin θ ′ n +1 cos( φ ′ n +1 − φ ′ n ) + cos θ ′ n cos θ ′ n +1 } + h { [ sinθ n sin θ n +1 × sin( φ n +1 − φ n ) − q 0 ] 2 + [sin θ ′ n sin θ ′ n +1 × sin( φ ′ n +1 − φ ′ n ) − q 0 ] 2 } + η { sin θ n sin θ ′ n × (cos φ n cos φ ′ n + sin φ n sin φ ′ n ) − cos θ n cos θ ′ n } + A (cos 2 θ n + cos 2 θ ′ n ) . (5) Using the equation of motion for the corresp onding quasi-spin mo del [19 ] in the limit A >> J, η , h , w e obtain ˙ φ n = 2 A cos θ n and ˙ φ ′ n = 2 A ′ cos θ ′ n . Hence, under absolute minima of 5 p oten tial the Hamiltonian (5 ) b e comes H = X n I 2 ( ˙ φ 2 n + ˙ φ ′ 2 n ) + J [2 − cos( φ n +1 − φ n ) − cos( φ ′ n +1 − φ ′ n )] − η [1 − cos( φ n − φ ′ n )] + h { 2 q 2 0 − [sin( φ n +1 − φ n ) − q 0 ] 2 − [sin( φ ′ n +1 − φ ′ n ) − q 0 ] 2 } , (6) where I = 1 2 A 2 is the momen t of inertia of the bases around the axes at p n ( p ′ n ). While rewriting the Hamiltonian in the a b ov e for m, w e ha v e restricted that the bases are rotating in the plane w hic h is normal to the helical axis. In otherw o rds , w e hav e now restricted our problem to a plane base rot a tor mo del [6] b y assuming θ = θ ′ = π / 2. Ha ving formed the Hamiltonia n, t he dynamics of the D NA double helix molecule can b e understo o d b y constructing the Hamilton’s equations of mo t io n corresp onding to the Hamiltonian (6) a s I ¨ φ n = [ J + 2 h cos( φ n +1 − φ n )] sin( φ n +1 − φ n ) − [ J + 2 h cos( φ n − φ n − 1 )] sin( φ n − φ n − 1 ) + η sin( φ n − φ ′ n ) − 2 hq 0 [cos( φ n +1 − φ n ) − cos( φ n − φ n − 1 )] , (7a) I ¨ φ ′ n = J + 2 h cos( φ ′ n +1 − φ ′ n ) sin( φ ′ n +1 − φ ′ n ) − J + 2 h cos( φ ′ n − φ ′ n − 1 ) sin( φ ′ n − φ ′ n − 1 ) + η sin( φ ′ n − φ n ) − 2 hq 0 [cos( φ ′ n +1 − φ ′ n ) − cos( φ ′ n − φ ′ n − 1 )] , (7b) where o verdot represen ts deriv ativ e with resp ectiv e to time. Eqs. (7a) and (7b) describ e the dynamics of the DNA double helix at the discrete lev el when the helical nature of the molecule is represen ted in the form of a t wist-lik e deformation. It is expected that the difference in angular rotation of bases with resp e ct to neighbouring bases along the t w o strands is small [5, 20]. Also, v ery recen tly Gaeta [8, 9] prop osed that the helical structure of DNA will in tr o duce q ualitativ e c hang es only in the small amplitude 6 regime. Hence, under the small a ngle approximation, in the con tinuum limit, the discrete equations of mot ion (7a,b) after suitable res caling of t ime and redefinition of the parameter η reduce to φ tt = ( J + 2 h ) I φ z z + η sin( φ − φ ′ ) , (8a) φ ′ tt = ( J + 2 h ) I φ ′ z z + η sin( φ ′ − φ ) . (8b) Adding a nd subtracting Eqs. (8a) and (8b) and after suitable rescaling of the v ariable z , w e obtain Ψ tt − Ψ z z + sin Ψ = 0 , (9) where Ψ = 2 φ a nd w e hav e further chos en 2 η = − 1. Also, while deriving Eq. (9), w e hav e c hosen φ ′ = − φ , b ecause among the p ossible rotations of bases, rotation of compleme n tary bases in opp osite direction easily facilitate an open state configuratio n. Eq. ( 9) is the com- pletely integrable sine-Go rdon equation whic h w as originally solv ed for N-soliton solutions in terms of kink and antik ink using the most celebrated Inv erse Scattering T ransform metho d [21]. F or instance, the kink and an tikink one soliton solution o f the sine-Gordon e quation is written in terms of the o r ig inal v ariables as φ ( z , t ) = 2 arctan[ exp [ ± 1 √ 1 − v 2 × s I ( J + 2 h ) ( z − v t )]] , (10) where + and − represe n t the kink and an tikink soliton solutions respectiv ely and v is t he v elo cit y o f the soliton. In Fig. 3(a ) w e hav e plotted the angular rotation of bases φ in terms of the kink-antikink one soliton solution a s given in Eq. (10) b y choosing the stac king, helicit y , momen t of inertia and v elo cit y parameters resp ectiv ely as J = 1 . 5 eV , h = 3 . 0 eV , I = 1 . 3 × 10 − 36 g cm 2 and v = 0 . 4 cm s − 1 [10, 1 5 ]. The kink-an tikink soliton solution whic h can propagate infinite distance and time describ es an op en state configuration in the DNA double helix whic h is sc hematically represen ted in Fig . 3(b). In order to understand the effect of helicity on the op en state configuration, in Fig. 3(c), w e hav e a ls o plott e d the kink and an tikink one soliton solution of the sine-Go r don equation in the absence of helicity , that is b y c ho osing h = 0 (k eeping all other parameters v alues the same), in the solution giv en 7 (a) (b) z t t z φ φ Kink Antikink Antikink Kink Base pair - 10 0 10 0 1 2 3 4 5 0 1 2 3 4 5 - 10 0 10 0 1 2 3 4 5 - 10 0 10 0 1 2 3 4 5 0 1 2 3 4 5 - 10 0 10 0 1 2 3 4 5 (c) z t t z φ φ (d) z Kink Antikink Kink Antikink Base pair - 10 0 10 0 1 2 3 4 5 0 1 2 3 4 5 - 10 0 10 0 1 2 3 4 5 - 10 0 10 0 1 2 3 4 5 0 1 2 3 4 5 - 10 0 10 0 1 2 3 4 5 FIG. 3: (a) Kink and an tikink one soliton solutio ns (Eq. (10)) of the sine-Gordon equation when helicit y is present ( h 6 = 0). (b) A sket c h of the formation of op en state configuration in terms of kink-an tikink solito ns in a DNA double h e lix when helicit y is present ( h 6 = 0). (c) Kink and an tikink one s o liton solutions of the sine-Go rdon equation when helicit y is absent (Eq. (10) wh e n h = 0). (d) A sk etc h of the formation of op en state configuration in terms of kink-an tikink solitons in DNA double h elix when helicit y is absent ( h = 0). 0 0.5 1 1.5 2 2.5 3 3.5 -8 -6 -4 -2 0 2 4 6 8 φ z h=0 h=1 h=2 h=3 h=4 0 0.5 1 1.5 2 2.5 0 1 2 3 4 5 Width Helicity (a) (b) FIG. 4: (a) T h e kink one soliton (Eq. (10)) represent ing base pair o p ening at t = 1 for different v alues of helicit y . (b) V ariation of wid th of the kink so liton against h eli cit y . in Eq.(10 ). F rom Figs. 3(a) and 3(c), w e observ e that when there is helicit y in the mo del ( h 6 = 0), the kink-an tikink soliton is getting broadened. In other w ords, helicit y in DNA mak es more num b er of base pairs to participate in the for ma t io n of op en stat e configuration without intro duc ing any qualitative c hange in the dynamics. This is a ls o sc hematically represen ted in Fig. 3(d) whic h lo oks eviden t on comparing Fig. 3(b). In order to highligh t the ab o v e fact, we hav e separately plotted the kink one solito n solution at a giv en time 8 ( t = 1) for differen t v alues of helicit y b y choosing h = 0 , 1 , 2 , 3 , 4 in F ig. 4(a). The increase in width aga inst helicity is explicitly represen ted in Fig . 4(b). F rom the figure it may b e noted that the increase in helicit y slo ws dow n the rotation of bases and mak es more and more n um b er of base pairs to part icipate in the op e n state configuration, th us pr oviding a b etter represen tation of base pair op ening in D NA. Th us, helicit y introduces a length scale v a riation in the base pair op ening. Similar results h a v e also b een o bserv ed b y Da ux ios [10] thro ugh a p erturbation a nalys is on his helicoidal mo del of DNA and obta ine d soliton with a muc h broader width. In order to ha v e a mor e realistic mo del, dissipativ e (viscous effect) and noise terms should b e added to the equations of motion. Experimentally , the life time of soliton in this case is shown to b e o f few nano seconds at ro om temp erature (see for e.g. Ref.[22]). Also, in a recen t pap er, Y akushevic h et al [23 ] through a n umerical analysis, sho w ed that when the viscosit y is strong the soliton mo v es a length o f only few c hain links and it will stop after that. On the other ha nd, when the viscosit y is low the soliton passes more than 3000 c hain links like a hea vy Bro wnian particle whic h is found to b e stable with resp ect to thermal oscillation. When the a bov e t w o effects a r e tak en in to accoun t Eq. ( 9 ) tak es the form Ψ tt − Ψ z z + sin Ψ = ǫ [ α Ψ t + β ( z , t )] where the terms prop ortional to α and β are related to viscous surrounding a nd thermal force s re sp ectiv ely . A solito n p erturbation ana lysis [24] on the ab o v e equation sho ws that when the viscosit y is high the soliton mov es for a small distance and then stops. But when the viscosit y is lo w the soliton mov es for a long distance along the c hain. The detailed analytical calculations of the ab o ve study will b e separately published elsewhe re. In summary , we prop osed a new helicoidal mo del to study D NA dynamics b y introducing the helical ch aracter in analogy with the twist deformation in a c holesteric liquid crystal system and the spin arrangemen t in a helimagnet. The nonlinear dynamics of DNA under the presen t helicoidal mo del is found to b e g ov erned by the completely inte grable sine- Gordon equation in the con tinu um limit whic h admits kink and an tikink soliton solutions. F ro m the nature of solitons, we observ e that helicit y in tro duces a length scale v aria tion without causing an y change in the shap e of the soliton. Due to this scaling v ariation, the width of the solito n increases and hence w e obtain bro a de r kink-an tikinks. In otherw ords a large num b er of bases are inv olv ed in the base pa ir op ening, th us leading to a b etter represen tation. This broadened base pair o pening may act as a b etter energetic activ ato r 9 in the case of RNA-P o lymerase transp ort during transcription pro cess in DNA. As the con tinuum helicoidal mo del do es not intro duc e qualitativ e c ha ng es in the DNA dynamics , w e prop ose to study the full nonlinear dynamics of the helicoidal model of DNA (without making small angle approx imation) b y solv ing equations (7a) and (7b) nume rically and the results will b e publishe d elsewhere. The w ork of M . D a nd V.V form pa rt of a ma jor D ST pro ject. [1] L. V. Y akushevic h, Nonlinear Physics of D NA (Wiley-V CH, Berlin, 2004). [2] S. W. Englander, N. R. Kallen b a nc h, A. J. Heeger, J. A. Krumh a nsl and S. Lit win, Proc. Natl. Acad. Sci. U .S.A 77 , 7222 ( 1980). [3] M. Peyrard and A. R. Bishop, Phys. Rev. Lett . 62 , 27 55 (1989). [4] V. Muto, P .S. Lomdahl and P .L. Christiansen, Phys. Rev. A 4 2 , 7452 (1990). [5] S. Y omosa, Phys. Rev. A 27 , 2120 (1983); 30 , 4 74 (1984). [6] S. T ak eno and S. Homma, Prog. T heor. Phys. 70 , 308 (1983); 7 2 , 679 (1984). [7] G. Gaeta, Ph ys. Lett. A 1 43 , 227 (1990); 168 , 383 (1 992). [8] G. Gaeta, Ph ys. Rev .E 74 , 0 21921 (2006). [9] G. Gaeta, J. Nonlin. Math. P hys. 14 , 5 7 (2007). [10] T. Dauxios, Phys. Lett. A 159 , 390 (1991). [11] M. Cadoni, R . De Leo and G. Gaeta, Ph ys. Rev. E 75 , 021919 (2007). [12] M. Barbi, S . Co cco and M. P eyrard, Phys. L ett. A 253 , 358 (1999). [13] M. Barbi, S . Lepri , M. Peyrard and N . Th e o dorak op oulos, Phys. Rev. E 68 , 0 61909 (2003). [14] A. Campa, Phys. R ev. E 63 , 021901 (2001). [15] S. T ak en o, Ph y s . Lett. A 339 , 3 52 (2005). [16] P . G. de Gennes, Physics of Liquid Crysta ls ( Clarendon Press, O xford) (1974). [17] R. M. White, Q uant um The ory of Magnetism (Sprin ger, New Y ork) (1982). [18] M. Daniel and J. Beula, Cho as Soliton and F r actals (Ac cepted for Publication) ( 2008). [19] J. Tjon and J . W right, Phys. Rev. B 15 , 3 470 (1977). [20] G. F a Zhou, Physica S c ripta 40 , 6 94 (1989). [21] M. J. Ablo witz, D. J. Kaup , A. C. New ell, a nd H. Segur, S tud. Appl. Math. 5 3 , 249 (1974). [22] M. P eyrard, N onlinearit y , 17 , R1 (2 004) and references therein. 10 [23] L. V. Y akushevic h, A. V. Sa vin a nd L. I. Manevitc h, Ph ys . Rev. E 66 , 016614 (2002). [24] M. Daniel and V. V asum athi, Ph ysica D 231 , 10 (2007). 11
Original Paper
Loading high-quality paper...
Comments & Academic Discussion
Loading comments...
Leave a Comment