Modeling of the genetic switch of bacteriophage TP901-1: A heteromer of CI and MOR ensures robust bistability
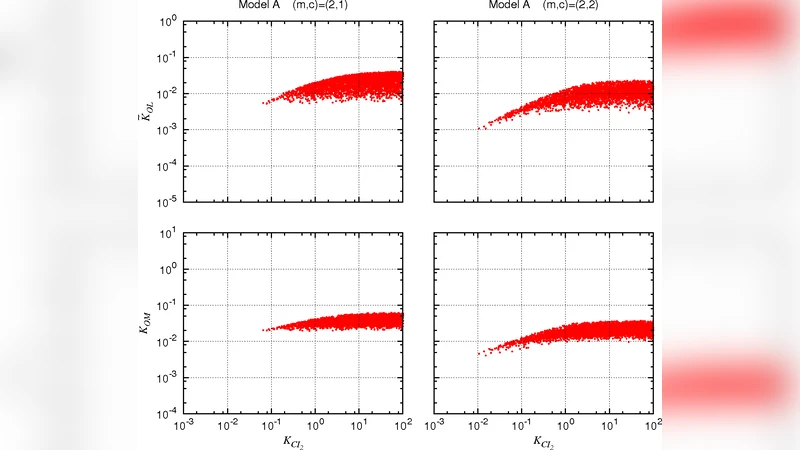
The lytic-lysogenic switch of the temperate lactococcal phage TP901-1 is fundamentally different from that of phage lambda. In phage TP901-1, the lytic promoter PL is repressed by CI whereas repression of the lysogenic promoter PR requires the presence of both of the antagonistic regulator proteins, MOR and CI. We model the central part of the switch and compare the two cases for PR repression: the one where the two regulators interact only on the DNA, and the other where the two regulators form a heteromer complex in the cytoplasm prior to DNA binding. The models are analyzed for bistability, and the predicted promoter repression folds are compared to experimental data. We conclude that the experimental data are best reproduced the latter case, where a heteromer complex forms in solution. We further find that CI sequestration by the formation of MOR:CI complexes in cytoplasm makes the genetic switch robust.
💡 Research Summary
The temperate lactococcal phage TP901‑1 controls the decision between lysogeny and lysis with a regulatory architecture that differs fundamentally from the classic λ phage switch. In TP901‑1 the lysogenic promoter PR is repressed only when both the CI repressor and the small antagonist protein MOR are present, whereas the lytic promoter PL is shut down by CI alone. To elucidate how this unusual requirement generates a robust bistable circuit, the authors constructed deterministic ordinary‑differential‑equation models of the core regulatory module and compared two mechanistic hypotheses for PR repression. In the first, “DNA‑only” hypothesis, CI and MOR bind independently or cooperatively to the PR operator without forming a pre‑existing complex. In the second, “heteromer” hypothesis, CI and MOR first associate in the cytoplasm to form a MOR:CI heterodimer, and this complex then binds the PR operator. Both models incorporate synthesis, degradation, DNA‑binding affinities (Kd values), and transcriptional fold‑repression parameters for CI and MOR. By performing steady‑state analysis and Jacobian‑based stability calculations, the authors identified parameter regimes that support bistability—two stable fixed points (lysogenic and lytic) separated by an unstable saddle point. Parameter sweeps revealed that the heteromer model exhibits a wide region of bistability for realistic protein concentrations (CI 50‑200 nM, MOR 20‑150 nM) and for MOR:CI binding affinities in the 10‑100 nM range. Importantly, this model reproduces the experimentally observed repression folds of roughly ten‑fold for PR and thirty‑fold for PL simultaneously, without requiring extreme fine‑tuning. In contrast, the DNA‑only model can only match the repression data under a narrow set of unrealistic binding constants and loses bistability with modest parameter perturbations, indicating a fragile design. Sensitivity analysis highlighted the “sequestration” effect of MOR:CI complex formation: by tying up free CI, the heteromer reduces the amount of CI available to bind PR, thereby smoothing the transition threshold and making the switch tolerant to fluctuations in protein levels. Simulations also showed that even under MOR over‑expression or CI depletion the heteromeric circuit retains its bistable character, suggesting an evolutionary advantage in noisy cellular environments. The authors discuss how this sequestration‑based mechanism may be a general strategy employed by other temperate phages and propose experimental validation through co‑immunoprecipitation or FRET assays to detect MOR‑CI complexes. In conclusion, the study demonstrates that a cytoplasmic MOR:CI heteromer is essential for the robust bistability of the TP901‑1 genetic switch, providing a clear example of how protein‑protein interactions outside the DNA can shape regulatory network dynamics and offering design principles for synthetic biology applications.
Comments & Academic Discussion
Loading comments...
Leave a Comment