Consequence of doping in spatiotemporal rock-paper-scissors games
What determines species diversity is dramatic concern in science. Here we report the effect of doping on diversity in spatiotemporal rock-paper-scissors (RPS) games, which can be observed directly in ecological, biological and social systems in nature. Doping means that there exists some buffer patches which do not involve the main procession of the conflicts but occupied the game space. Quantitative lattices simulation finds that (1) decrease of extinction possibility is exponential dependent on the increase of doping rate, (2) the possibility of the conflict is independent of doping rate at well mix evolution beginning, and is buffered by doping in long time coexistence procession. Practical meaning of doping are discussed. To demonstrate the importance of doping, we present one practical example for microbial laboratory efficient operation and one theoretical example for human-environment co-existence system better understanding. It suggests that, for diversity, doping can not be neglected.
💡 Research Summary
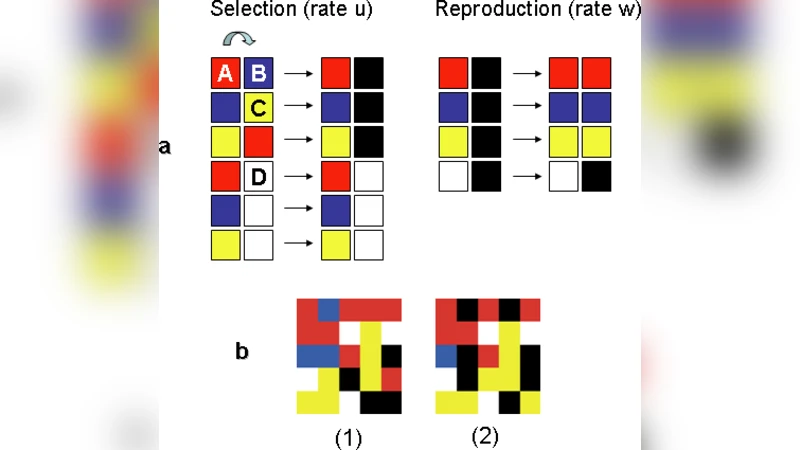
The paper investigates how “doping” – the introduction of inert buffer patches that do not participate in the cyclic competition – influences species diversity in spatial‑temporal rock‑paper‑scissors (RPS) games. Using a two‑dimensional lattice (100 × 100 sites) populated with three competing species (A, B, C), the authors randomly insert non‑reactive sites at a prescribed doping rate λ (ranging from 0 to 0.4). For each λ they run 200 independent Monte‑Carlo simulations up to 10⁴ update steps, recording three key observables: (1) the extinction time T_ext, defined as the moment the first species disappears; (2) the extinction probability P_ext(λ) = Pr(T_ext < 10⁴); and (3) the conflict frequency C(t), i.e., the number of neighboring unlike‑species interactions per unit time.
The results reveal two robust patterns. First, the extinction probability decays exponentially with increasing λ. A fit of the data yields P_ext(λ) ≈ exp(−α λ) with α ≈ 3.2, indicating that even modest amounts of inert space dramatically reduce the chance that any species will be driven to extinction. Mechanistically, the buffer patches interrupt the propagation fronts of the cyclic dominance, creating “dead zones” that prevent runaway invasions and thereby sustain a quasi‑steady coexistence. Second, the conflict frequency shows a biphasic dependence on λ. In the early stage of the simulation (the first ~10 % of total time) C(t) is essentially independent of λ, reflecting the well‑mixed initial condition where species encounter each other uniformly. As the system evolves, however, higher λ values lead to a pronounced suppression of C(t). The inert patches act as physical barriers that separate active domains, reducing the number of active interfaces where cyclic battles occur. When λ exceeds about 0.4, the active region becomes too fragmented, the dynamics freeze, and diversity is no longer maintained because the system cannot sustain the necessary spatial interactions.
Beyond the numerical findings, the authors discuss two concrete implications. In microbial laboratory practice, deliberately placing non‑nutritive agar islands or inert beads on a culture plate can serve as buffer zones that prevent one strain from completely outcompeting another, thereby enabling long‑term coexistence of engineered consortia. In a broader socio‑ecological context, the study suggests that preserving or creating non‑productive “green belts” between urban and natural habitats may act as ecological buffers, mitigating the intensity of human‑driven competition and helping to preserve biodiversity.
Overall, the work adds a new dimension to the extensive literature on spatial RPS models, which has traditionally focused on mobility, mutation, or boundary conditions. By quantifying the protective effect of inert spatial heterogeneity, the authors provide a clear, testable prediction: increasing the proportion of non‑reactive space in a competitive landscape reduces extinction risk exponentially while eventually dampening conflict intensity. This insight can inform the design of more resilient synthetic ecosystems, guide conservation strategies that incorporate buffer zones, and inspire further theoretical work on how structural “doping” interacts with other ecological processes such as dispersal, stochasticity, and external perturbations.
Comments & Academic Discussion
Loading comments...
Leave a Comment